╚ń║╬░čėąÖCÅU╦«▐D╗»×ķŪÕØŹ─▄į┤
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2016-10-16 8:30:13
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ1 ę²čį
ĪĪĪĪę└┘ćė┌╬ó╔·╬’Ą─ģÆ觎¹╗»ū„ė├─▄ē“īóČÓĘNėąÖCÅU╦«║═╣╠¾wÅU╬’▐D╗»×ķ╝ū═ķĪóÜõÜŌĄ╚ŪÕØŹ─▄į┤Ż¼╩Ūę╗ĘNŁhŠ│ėč║├ą═Ą─ą┬─▄į┤╔·«a═ŠÅĮŻ¼ę▓╩Ū─┐Ū░╠Ä└Ē╣ż▐rśIėąÖCÅU╬’Ą─ę╗éĆųžę¬╩ųČ╬.ģÆ觎¹╗»Ą─║╦ą─╩Ū╬ó╔·╬’╚║Ż¼ę“┤╦Ż¼½@Ą├Ė▀╗ŅąįĄ─ģÆ觎¹╗»Š·╚║╩Ūėąą¦▐D╗»ėąÖCÅUŚē╬’×ķ─▄į┤╬’┘|Ą─ųžę¬▒ŻšŽ.╔·╬’Ę┤æ¬Ų„ā╚Ą─ģÆ觎¹╗»Š·╚║Š▀ėąÅ═ļsĄ─ĮM│╔Ż¼═©│Ż▒╗äØĘų×ķ3ŅÉŻ¼╝┤╦ß╗»Š·╚║Īó«aÜõ«aęę╦ߊ·╚║ęį╝░«a╝ū═ķŠ·╚║.Ųõ╗Ņąį╩▄ČÓĘNę“╦žė░ĒæŻ¼░³└©Ę┤æ¬Ų„Ą─ĮYśŗĪópHųĄĪó╦«┴”═Ż┴¶ĢrķgĄ╚.NadarajahĄ╚šJ×ķ£žČ╚╩ŪģÆ觎¹╗»Š·╚║Ą─ųžę¬ė░Ēæę“ūėŻ¼īóė░Ēæ╔·╬’Ę┤æ¬Ų„ā╚Ą─Š·╚║ĮYśŗĪó╗»īWąĶč§┴┐(Chemical Oxygen DemandŻ¼COD)╚ź│²┬╩Īó│÷╦«æęĖĪ╣╠¾w(Effluent Suspended SolidsŻ¼ESS)║═╬█─ÓųĖöĄ(Sludge Volume IndexŻ¼SVI)Ą╚.
ĪĪĪĪ═©│ŻģÆ觎¹╗»ū„ė├į┌2éĆ£žČ╚ĘČć·ā╚▀MąąŻ¼╝┤ųą£žģÆ觎¹╗»║═Ė▀£žģÆ觎¹╗».┴╝║├Ė▀£žģÆ觎¹╗»Š·╚║į┌±Z╗»ų«│§Ż¼Š·į┤│Ż╚Īūįė┌ūį╚╗ŁhŠ│╚ń═┴╚└Īó║ėĄ└Ąū─ÓĪó┼Żč“╝S▒Ńęį╝░│Ż£ž╗Ņąį╬█─ÓĄ╚.į┌╠ß╔²£žČ╚Ą─Ė╗╝»┼ÓB▀^│╠ųąŻ¼ųØuą╬│╔ĘĆČ©Ą─ģÆ觎¹╗»Š·╚║.PetterssonĄ╚Ą─蹊┐šJ×ķŻ¼┼ÓB£žČ╚Ė─ūāĢ■ĮoŠ·╚║ĮYśŗĄ─š{š¹Ä¦üĒą┬Ą─▀xō±ē║┴”.£žČ╚▐DūāĘ∙Č╚įĮ┤¾Ż¼▀@ĘN▀xō±ē║┴”įĮÅŖ┴ęŻ¼▓óį÷╝ėŠ·╚║ĮYśŗš{š¹Ą─Ę∙Č╚.═¼ĢrŻ¼£žČ╚┐╔─▄ė░Ē抷╚║ūā╗»Ą─ų▄▐D╦┘┬╩Ż¼Ė▀Ą─ų▄▐D╦┘┬╩╩╣Š·╚║Ė³╝ė┐ņ╦┘Ą─Ė³ōQĄ¶▓╗▀mæ¬ą┬£žČ╚Ą─Š·╚║.ZhengĄ╚└¹ė├Ęųūė╔·╬’īWĄ─Öz£yĮY╣¹░l¼FŻ¼Ė▀£žĄ─«a╝ū═ķ╣┼Š·┐╔┤µį┌ė┌│Ż£žģÆč§Ę┤æ¬Ų„ųąŻ¼Ę┤ų«ęÓ╚╗Ż¼Č°Ųõ╔·╬’┴┐╚ĪøQė┌┼ÓB£žČ╚.Ahring╠Į╦„┴╦Ė▀£žģÆ觎¹╗»▀^│╠£žČ╚ūā╗»ī”Š·╚║╔·╬’┴┐Ą─ė░ĒæŻ¼ĮY╣¹’@╩Š«öīóę╗éĆ▀B└möć░ĶĘ┤æ¬Ų„Ą─┼ÓB£žČ╚Å─55 Īµ╠ßĖ▀ĄĮ65 ĪµŻ¼╝ÜŠ·Ą─╔·╬’┴┐īóėą├„’@Ą─Ž┬ĮĄŻ¼Č°╣┼Š·Ą─╔·╬’┴┐Ģ■ėą╔Ž╔²Ż¼«ö£žČ╚▀_ĄĮ65 ĪµĢrŻ¼ÜõĀIBą═«a╝ū═ķ╣┼Š·Š▀ėą├„’@Ą─╔·╬’╗Ņąį.
ĪĪĪĪ┐╔ęŖŻ¼£žČ╚Ą─▐Dūāī”ģÆ觎¹╗»Š·╚║ĮYśŗ║═╣”─▄Č╝ėąÅŖ┴ęĄ─ė░Ēæ.×ķ┴╦£p╔┘£žČ╚ī”ģÆ觎¹╗»Š·╚║Ą─ø_ō¶Ż¼į┌ģÆč§╔·╬’Ę┤æ¬Ų„åóäėĢr═©│Ż▓╔ė├╠▌Č╚╔²£žĄ─ĘĮĘ©(Ēnė²║ĻĄ╚Ż¼2009).▒ŠčąŠ┐ät└¹ė├37 ĪµĄ─ųą£žģÆ觎¹╗»Š·╚║ų▒Įė▐D╚ļ50 Īµ┼ÓBŻ¼▓ó═©▀^▀B└m▀M┴Ž╠Ū├█ėąÖCÅU╦«±Z╗»Ė▀£žģÆ觎¹╗»Š·╚║Ż¼▀MČ°┐╝▓ņĖ▀£ž±Z╗»▀^│╠ųąŠ·╚║ĮYśŗĪóČÓśėąį╝░╣”─▄Ą─ūā╗»╠žš„.蹊┐ĮY╣¹ī”ĻU├„ģÆč§░lĮ═Š·╚║į┌£žČ╚äĪ┴ęūā╗»Ä¦üĒĄ─▀xō±ē║┴”Ž┬Š·╚║ĮYśŗ┼c╣”─▄Ą─ūā╗»┌ģä▌Ż¼ī”Ė─▀MĖ▀£žģÆ觎¹╗»Š·╚║Ė╗╝»ĘĮĘ©Š▀ėąųžę¬ęŌ┴x.
ĪĪĪĪ2 ▓─┴Ž┼cĘĮĘ©
ĪĪĪĪ2.1 ģÆč§Ę┤æ¬Ų„▀\ąąŚl╝■
ĪĪĪĪ400 mLīŹ“×╩ę┼ÓBĄ─ųą£ž(37 Īµ)ģÆ觎¹╗»ĮšČÆĪóžiĪó┼Ż╝S▒ŃĄ─╗Ņąį╬█─Óūó╚ļ1 LūįųŲģÆč§Ę┤æ¬Ų„ųąŻ¼ūįüĒ╦«ča│õų┴800 mLŻ¼Ę┼ų├ė┌50 ĪµŚl╝■Ž┬┼ÓB.Ė▀ØŌČ╚╠Ū├█ėąÖCÅU╦«(100000 mgĪżL-1 COD)(└ŅĮ▄Ą╚Ż¼2011)ū„×ķģÆ觎¹╗»įŁ┴ŽŻ¼├┐2 d▀M┴Ž1┤╬Ż¼├┐┤╬30 mL(╦«┴”═Ż┴¶Ģrķg(Hydraulic Retention TimeŻ¼HRT)=27 d).├┐┤╬▀M┴ŽŪ░Å─Ę┤æ¬čbų├ųą│ķ│÷30 mLģÆ觎¹╗»ę║Ż¼ė├ė┌└Ē╗»ųĖś╦┼c╬ó╔·╬’ČÓśėąįĘų╬÷Ż¼ęį╝░ŠS│ųĘ┤款wŽĄ¾wĘeŲĮ║ŌŻ¼ģÆč§Ę┤æ¬Ų„čbų├ęŖłD 1.
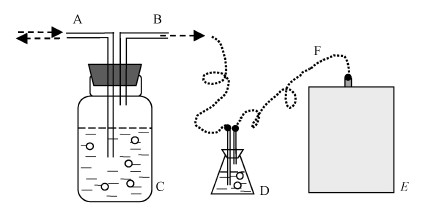
ĪĪĪĪłD 1ģÆ觎¹╗»Ę┤æ¬Ų„(AŻ║▀M/╚Ī┴Ž┐┌, BŻ║│÷ÜŌ┐┌, CŻ║ģÆč§░lĮ═Ų┐, DŻ║ŠÅø_čbų├, EŻ║╝»ÜŌ┤³, FŻ║▀BĮė╣▄)
ĪĪĪĪ2.2 └Ē╗»ųĖś╦Ęų╬÷
ĪĪĪĪ1)pH ╚Ī┤²£yę║0.5 mLŻ¼ė├╚š▒ŠHORIBA B-212ą═╬ó┴┐pHėŗ(Compact pH meterŻ¼model B-212Ż¼Japan)£yČ©pHųĄ.
ĪĪĪĪ2)╗»īWąĶč§┴┐(COD)0.25 mmolĪżL-1 K2Cr2O7 ╚▄ę║┼cAg2SO4 ╚▄ę║(Ag2SO4 /ØŌH2SO4┘|┴┐¾wĘe▒╚=1/100)░┤¾wĘe▒╚1:3╗ņ║ŽŻ¼ėø×ķ╚▄ę║A.╚Ī2 mL ┤²£yśėŲĘĪó3 mL ╚▄ę║AĪó╔┘įSHgSO4(Ų┴▒╬Cl-)Ż¼╝ė╚ļ10 mL Ž¹ĮŌ╣▄ųąŻ¼į┌Ž¹ĮŌāx╔ŽŽ¹ĮŌ(150 Īµ)2 h.Ž¹ĮŌĄ─śėŲĘ└õģs║¾└¹ė├Ęų╣Ō╣ŌČ╚ėŗ£yČ©śėŲĘODųĄ.┐š░ū┐žųŲ╩╣ė├š¶s╦«╠µ┤·śėŲĘ╚▄ę║.
ĪĪĪĪ3)šėÜŌ┐é┴┐╝░╝ū═ķ║¼┴┐ ├┐╚š╔·«ašėÜŌ╩╣ė├╝»ÜŌ┤³╩š╝»Ż¼└¹ė├┼┼╦«╝»ÜŌĘ©£yČ©┐é┴┐.šėÜŌųą╝ū═ķ║¼┴┐ė├ėóć°Geotech ╣½╦ŠĄ─šėÜŌ│╔Ęų£yČ©āx(ą═╠¢×ķBiogas Check)£yČ©.
ĪĪĪĪ4)┐é╣╠¾w(Total SolidsŻ¼TS)║¼┴┐║═ō]░ląį╣╠¾w(Volatile SolidŻ¼VS)║¼┴┐ ▓╔ė├ųž┴┐Ę©£yČ©.
ĪĪĪĪ5)ō]░ląįėąÖC╦ß(Volatile Fatty AcidŻ¼VFA)║¼┴┐ 1 mLŽ¹╗»ę║10000 rĪżmin-1ļxą─15 minŻ¼╚Ī0.5 mL╔ŽŪÕŻ¼╝ė╚ļ0.5 mLęęŪĶŻ¼│õĘų╗ņä“║¾Ż¼▒∙╔Ž▒Ż│ų10 minŻ¼10000 rĪżmin-1ļxą─15 min║¾▀^0.22 ”╠m╚ź│²Ą░░ū.Ū░╠Ä└ĒśėŲĘÖz£yėąÖC╦߯¼╩╣ė├LC/MS-2010AĖ▀ą¦ę║ŽÓ╔½ūV(ShimadzuŻ¼KyotoŻ¼Japan).Öz£yŚl╝■×ķŻ║10 ”╠LśėŲĘūó╚ļāxŲ„Öz£yŻ¼Öz£yų∙×ķAminex HPX-87H Column(Bio-RadŻ¼CAŻ¼USA)Ż¼0.05 mmolĪżL-1 H2SO4×ķ┴„äėŽÓ.Ž┤├ō│╠ą“×ķ0.6 mLĪżmin-1 5 minŻ¼0.4 mLĪżmin-1 30 minŻ¼0.6 mLĪżmin-1 5 min.ų∙£ž40 ĪµŻ¼Öz£yŲ„×ķČ■śO╣▄Ļć┴ąÖz£yŲ„.
ĪĪĪĪ2.3 ╬ó╔·╬’ČÓśėąįĘų╬÷
ĪĪĪĪ1)DNA╠ß╚Ī╝░PCRŚl╝■ ▒∙ā÷▒Ż┤µśėŲĘ└¹ė├ė┘─Ó╗∙ę“ĮMDNA┐ņ╦┘╠ß╚Īįćä®║ą(BioTekeŻ¼░┘╠®┐╦╔·╬’╝╝ągėąŽ▐╣½╦ŠŻ¼▒▒Š®)╠ß╚Ī┐éDNAŻ¼ĘĮĘ©░┤šf├„.│ķ╠ߥ─┐éDNA└¹ė├ę²╬’▀MąąöUį÷Ż¼Ę┤款wŽĄ(50 ”╠L):─Ż░ÕDNA 10 ngŻ¼10 Ī┴PCR Gold Buffer 5 ”╠LŻ¼25 mmolĪżL-1 MgCl2Ż¼3 ”╠LŻ¼dNTP mix(Ė„2 mmolĪżL-1)5 ”╠LŻ¼45 ”╠molĪżL-1 ę²╬’(27f║═906r)Ė„0. 5 ”╠LŻ¼5 Unit Ampli Taq Gold 0. 2 ”╠L.PCRĘ┤æ¬Śl╝■×ķŻ║94 ĪµŅAūāąį10 minŻ¼94 Īµūāąį30 sŻ¼52 Īµ═╦╗45 sŻ¼72 Īµčė╔ņ1 min 30 sŻ¼╣▓22éĆ裣hŻ¼ūŅ║¾į┌72 ĪµŽ┬čė╔ņ5 min.PCR«a╬’ė├15 mgĪżmL-1Ą─Łéų¼╠Ū─²─zļŖėŠÖz£y.
ĪĪĪĪ2)ūāąį╠▌Č╚─²─zļŖėŠ(DGGE)╠ß╚ĪģÆ觎¹╗»Š·╚║DNAŻ¼└¹ė├PCRāxöUį÷Š·╚║16S rDNA╗∙ę“Ų¼Č╬Ų¼Č╬(╩╣ė├ę²╬’Ż¼╝ÜŠ·Ż║357F-GCŻ¼517 R;╣┼Š·Ż║A348IFŻ¼U806IR-GC).╩╣ė├DcodeTM Universal Mutation Detector System(BIO-RAD LaboratoriesŻ¼HerculesŻ¼CAŻ¼USA)ī”╦∙½@DNAŲ¼Č╬▀Mąą─²─zļŖėŠ.ūāąį─z1 mm ║±Ż¼PolyacrylamideØŌČ╚╠▌Č╚6%~12%(W/V)Ż¼0.5Ī┴ TAB Electrophoresis Buffer(20 mmolĪżL-1 Tris-HClŻ¼pHųĄ 8.3Ż¼10 mmolĪżL-1 acetic acidŻ¼0.5 mmolĪżL-1 EDTA)Ż¼ūāąįä®─“╦ž╠▌Č╚20%~55%(║¼ėą40%╝ū§Ż░ĘŻ¼7 molĪżL-1─“╦ž).ļŖėŠļŖē║200 VŻ¼£žČ╚×ķ61 ĪµŻ¼Ģrķg5 h.ļŖėŠĮY╩°║¾ė├SYBR Green I(Molecular ProbesŻ¼EugeneŻ¼OR.)╚Š╔½Ż¼ė^▓ņ║═ššŽÓ╩╣ė├Alpha Innotech─²─z│╔Ž±āx.Ė∙ō■ļŖėŠŚlĦŻ¼Ęų╬÷Š·╚║ĮYśŗĘĆČ©ąį.
ĪĪĪĪ3)┐╦┬Ī╬─Äņ īó╝ā╗»║¾Ą─PCR«a╬’(╩╣ė├ę²╬’Ż¼╝ÜŠ·Ż║27FŻ¼907R;╣┼Š·Ż║109FŻ¼912R)▀BĮėĄĮpGEM-T Easy▌d¾wŻ¼4 Īµ▀^ę╣Ż¼╚╗║¾═©▀^į┌42 Īµ ¤ß╝ż 45 s▐D╗»ĄĮE.coli JM109Š·¾wā╚;═©▀^X-Gal║═IPTGĄ─”┴-╗źčaū„ė├║Y▀x│÷░ū╔½Š·┬õ.▓óīó░ū╔½Š·┬õ▐DęŲĄĮą┬§rĄ─║¼ėą┐╣╔·╦žĄ─LB╣╠¾w┼ÓB╗∙╔ŽŻ¼ŠÄ┤aĮ©┴ó┐╦┬Ī╬─Äņ.└¹ė├ūāąį╠▌Č╚─²─zļŖėŠ(Denaturing Gradient Gel EletrophoresisŻ¼DGGEŻ¼ūāąįä®ØŌČ╚ĘČć·įO×ķ20%~60%)║Y▀x│÷▓╗═¼ĘNŅÉĄ─┐╦┬Īūė.ī”║¼ėą─┐ś╦┐╦┬ĪūėĄ─Š·¾wŻ¼ė├ēA┴čĮŌĘ©╠ß╚Ī┘|┴ŻŻ¼▓ó£yą“.└¹ė├GenBank(NCBIŻ¼▀MąąŽÓ╦ŲŠ·ųĻÖz╦„Ż¼½@╚ĪĮ³ŠēŠ·ųĻŽÓĻPą“┴ą.æ¬ė├DNAą“┴ąĘų╬÷▄ø╝■DNA Star 7.0║═MEGA 5.0▄ø╝■▀Mąąą“┴ąŲ┤ĮėŻ¼śŗĮ©ŽĄĮy░lė²śõ(NJĘ©).
ĪĪĪĪ4)Č©┴┐PCR ╣┼Š·┐é┴┐Č©┴┐PCR▓╔ė├Platinum ® SYBR ® Green qPCR SuperMIx-UDGįćä®║ą.Ę┤款wŽĄ(20”╠L)×ķŻ║2Ī┴Platinum SYBR ® Green qPCR SuperMix-UDGŻ¼10 ”╠L;50 mmolĪżL-1 MgCl2Ż¼1.2 ”╠L;Forward and Reverse specific primers(10 pmolĪż”╠L-1)Ż¼Ė„0.4 ”╠L;DNAŻ¼1 ”╠L;RoxŻ¼0.4 ”╠L;PCR grade distilled water 7.8 ”╠L.ś╦£╩Ū·ŠĆųŲū„Ż¼╩╣ė├║¼ėą▒╗Öz£yŠ·ųĻ─┐ś╦Ų¼Č╬┐╦┬ĪūėĄ─pGEM-T Easy┘|┴Żū„×ķś╦£╩śėŲĘŻ¼ė├¤╔╣ŌČ©┴┐āxŠ½┤_£yČ©ś╦£╩śėŲĘØŌČ╚Ż¼▓óėŗ╦ŃŲõ┐ĮžÉöĄ.īóŽĪßī│╔▓╗═¼öĄ┴┐╝ē┐ĮžÉöĄĄ─╠▌Č╚ś╦£╩ę║Ż¼┼c▒╗£yśėŲĘę╗═¼╩╣ė├ABI Prism ® 7500ą═¤╔╣ŌČ©┴┐PCRāx(ABIŻ¼USA)▀MąąPCR╝░¤╔╣ŌÖz£y.öUį÷│╠ą“▓╔ė├ABIāxŲ„Ą─ś╦£╩│╠ą“Ż║50 Īµ│ų└m 2 min(UDGʧė²)Ī· 95 Īµ│ų└m2 min Ī· 40éĆ裣hŻ║95 ĪµŻ¼15 s;60 ĪµŻ¼30 s.¤╔╣ŌÖz£yŲ„į┌60 ĪµĢr£yČ©¾wŽĄ¤╔╣ŌÅŖČ╚.öĄō■Ęų╬÷Ż¼═©▀^CtųĄ┤_Č©ś╦£╩Ū·ŠĆŻ¼įuāröĄųĄĄ─ā×┴╝Ż¼ūŅĮK½@╚Ī┤²£yśėŲĘ┐ĮžÉöĄ(╔·╬’┴┐).Methanomicrobiales(MMB)Ż¼Methanococcales(MCC)Ż¼Methanosaetaceae(MST)Ęų╬÷╩╣ė├TaqMan ® Universal PCR Master Mix(Applied BiosystemsŻ¼USA)įćä®║ąĘų╬÷Ż¼Ę┤款wŽĄ×ķPCR-grade waterŻ¼5 ”╠L;Forward and Ż¼Ė„1 ”╠L;ī”æ¬TaqMan╠Įßś(1 ”╠molĪżL-1)1 uL;2Ī┴TaqMan ® Universal PCR Master MixŻ¼10 ”╠L; DNAŻ¼1 ”╠L.Ųõ╦¹═¼Platinum ® SYBR ® Green qPCR SuperMIx-UDGįćä®║ąĘ┤æ¬.ŲõųąMMB═╦╗£žČ╚×ķ63 Īµ.
ĪĪĪĪ3 ĮY╣¹
ĪĪĪĪ3.1 Ė▀£ž±Z╗»▀^│╠ųąCODĪópHųĄ║═Š·╚║«ašėÜŌ┴┐Īó╝ū═ķ║¼┴┐ūā╗»
ĪĪĪĪīóüĒūįė┌īŹ“×╩ęĄ─37 ĪµŚl╝■Ž┬▀\ąąĄ─Ū▌ą¾╝S▒ŃģÆ觎¹╗»«a╝ū═ķŠ·╚║ų▒Įė▐DęŲų┴50 ĪµŚl╝■Ž┬┼ÓBŻ¼╠Ä└ĒĖ▀ØŌČ╚ėąÖCÅU╦«.Ė▀£ž±Z╗»Ų┌ķgŻ¼ģÆ觎¹╗»Š·╚║Ą─«aÜŌ┴┐Īó╝ū═ķ║¼┴┐ĪópHųĄ╝░CODĄ╚└Ē╗»╠žš„ųĖś╦▒╗Öz£y(łD 2).ĮY╣¹▒Ē├„ųą£žĄ─ģÆ觫a╝ū═ķŠ·╚║▐D╚ļĖ▀£ž┼ÓBĄ─Ū░7 d╝┤ķ_╩╝«aÜŌŻ¼ŲĮŠ∙╚š«aÜŌ┴┐×ķ115 mLĪżd-1Ż¼20 d║¾Ż¼«aÜŌ┴┐▀M╚ļĖ▀ĘÕŲ┌Ż¼ŲĮŠ∙╚š«aÜŌ┴┐│¼▀^360 mLĪżd-1.ūŅĖ▀╚š«aÜŌ┴┐│÷¼Fį┌┼ÓBĄ─Ą┌31 dŻ¼▀_ĄĮ410 mLĪżd-1(łD 2d).┼ÓBŲ┌ķg┐éėŗ«aÜŌ┴┐×ķ9735 mL(łD 2d)Ż¼ŲĮŠ∙COD▐D╗»ą¦┬╩×ķ162.5 mLĪżg-1.╝ū═ķ║¼┴┐Ą─ūā╗»┌ģä▌┼cŲĮŠ∙╚š«aÜŌ┴┐ŽÓ╦ŲŻ¼│§Ų┌╝ū═ķ║¼┴┐▌^Ą═(18.9%)Ż¼20 d║¾Ż¼╝ū═ķ║¼┴┐ŠS│ųį┌34%ęį╔Ž.Ę┤æ¬Ų„ā╚Ą─COD║¼┴┐į┌«aÜŌŲ┌ķgĘĆČ©į┌6800~7700 mgĪżL-1(łD 2a)Ż¼pHųĄŠS│ųį┌7.3~8.1ų«ķg(łD 2b).4éĆģóöĄĄ─Öz£yĮY╣¹▒Ē├„ģÆ觎¹╗»Š·╚║▒╗▐Dų┴Ė▀£žŚl╝■Ž┬±Z╗»┼ÓBŻ¼─▄ē“┐ņ╦┘åóäėŻ¼▓ó▀B└mĪóĘĆČ©Ąž╠Ä└ĒĖ▀ØŌČ╚ėąÖCÅU╦««a╔·šėÜŌ.
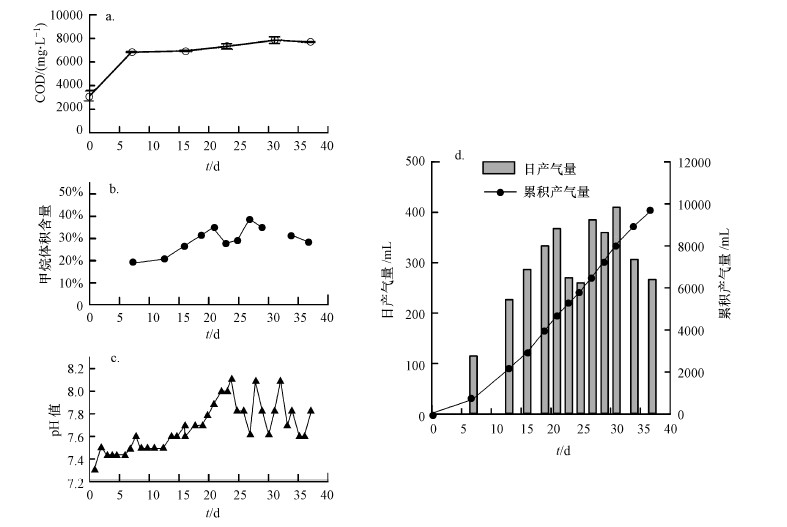
ĪĪĪĪłD 2Ė▀£ž±Z╗»ģÆ觎¹╗»Š·▀^│╠ųąCODĪópHųĄ║═Š·╚║«ašėÜŌ┴┐Īó╝ū═ķ║¼┴┐ūā╗»(ūóŻ║▒ŠčąŠ┐ųą0 dśėŲĘ×ķ30 Īµ┼ÓBĄ─įŁ╩╝ģÆ觎¹╗»ę║Ż¼Ųõ╦¹Ģrķg×ķ▐DęŲų┴50 Īµ║¾Ą─ģÆ觎¹╗»ę║śėŲĘŻ¼Ž┬═¼.)
ĪĪĪĪ3.2 Ė▀£ž±Z╗»ģÆ觎¹╗»Š·╚║▀^│╠ųąėąÖC╦ß║¼┴┐ūā╗»
ĪĪĪĪ▒¹╦ß║═ęę╦ß╩Ū░lĮ═ę║ųą║¼┴┐ūŅĖ▀Ą─ā╔ŅÉėąÖC╦ß(łD 3).▒¹╦ß║¼┴┐į┌åóäėŪ░Ų┌š╝ėą├„’@Ą─ā×ä▌Ż¼ūŅĖ▀ųĄ×ķ784.7 mgĪżL-1(Ą┌15 d)Ż¼«öĘ┤æ¬Ų„▀\ąąĀŅæBĘĆČ©║¾Ż¼▒¹╦ß║¼┴┐’@ų°Ž┬ĮĄŻ¼į┌┼ÓBĄ─Ą┌30 dŻ¼ĮĄĄ═ų┴25.3 mgĪżL-1.ęę╦ß╩Ū«a╝ū═ķ╣┼Š·║Ž│╔╝ū═ķĄ─ųžę¬Ąū╬’Ż¼Ę┤æ¬Ų„ā╚ęę╦ߥ─Ęe└█╝░▐D╗»╦┘Č╚│Ż│ŻĘ┤æ¬┴╦ģÆ觎¹╗»Š·╚║╔·«a╝ū═ķĄ─ĀŅæB.į┌Ę┤æ¬Ų„åóäėŪ░Ų┌Ż¼░lĮ═ę║ā╚│÷¼Fęę╦ßĘe└█¼FŽ¾Ż¼▓óļS┼ÓBĢrķgĄ─į÷╝ėČ°į÷╝ėŻ¼į┌Ą┌15 d║¼┴┐▀_ĄĮ439.3 mgĪżL-1.▀M╚ļ«aÜŌĖ▀ĘÕ║¾Ż¼Ę┤款wŽĄā╚Ą─ęę╦ß║¼┴┐├„’@ĮĄĄ═Ż¼į┌┼ÓBĄ─Ą┌30 dŻ¼ęę╦ß║¼┴┐£p╔┘ų┴145.3 mgĪżL-1.╝ū╦ß║═╚ķ╦ßį┌░lĮ═ę║ųąĄ─║¼┴┐▌^╔┘Ż¼Ųõųą╚ķ╦ß┘|┴┐ØŌČ╚18~81 mgĪżL-1ų«ķgŻ¼╝ū╦ßāHį┌░lĮ═Ū░Ų┌▒╗Öz£yĄĮŻ¼┘|┴┐ØŌČ╚╝sį┌16~18 mgĪżL-1ų«ķg.
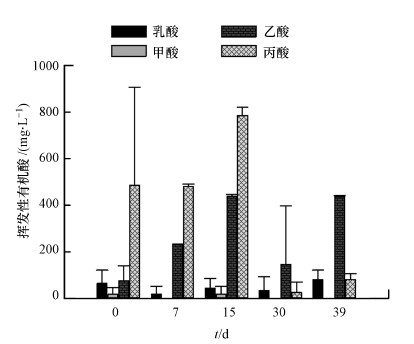
ĪĪĪĪłD 3ģÆ觎¹╗»▀^│╠ėąÖC╦ß║¼┴┐ūā╗»
ĪĪĪĪ3.3 Ė▀£ž±Z╗»ģÆ觎¹╗»Š·╚║▀^│╠ųąŠ·╚║ĮYśŗĘų╬÷
ĪĪĪĪĘ┤æ¬Ų„▀\ąąŲ┌ķgŻ¼ģÆ觎¹╗»Š·╚║ųą╝ÜŠ·║═╣┼Š·Ą─Š·╚║ĮYśŗ└¹ė├DGGE╝╝ąg▀MąąĘų╬÷.ĮY╣¹’@╩ŠŻ¼į┌╝ÜŠ·Š·╚║Ą─DGGEłDūVųąŻ¼┤¾┴┐Ą─Ė▀┴┴ŚlĦ┤µį┌ė┌├┐éĆėŠĄ└ā╚Ż¼▒Ē├„╝ÜŠ·Š·╚║Ą─ČÓśėąį╩«ĘųžSĖ╗(łD 4aĪó4b)Ż¼Č°╣┼Š·Š·╚║Ą─Ė▀┴┴ŚlĦŽÓī”ė┌╝ÜŠ·Š·╚║├„’@£p╔┘.į┌Ę┤æ¬Ų„▀\ąąŲ┌ķgŻ¼¤ošō╝ÜŠ·▀Ć╩Ū╣┼Š·Š·╚║Ż¼Č╝┤µį┌ę╗ą®ŚlĦĄ─Ž¹╩¦║═│÷¼FĄ─ĀŅør.į┌╝ÜŠ·Š·╚║ųą╝sėą7éĆų„ꬌlĦ│÷¼F(Ī░Ī„Ī▒ųĖ╩Š▓┐ĘųŻ¼Ž┬═¼)Ż¼7éĆų„ꬌlĦŽ¹╩¦(Ī░![]() Ī▒ųĖ╩Š▓┐ĘųŻ¼Ž┬═¼);╣┼Š·Š·╚║ųą╝sėą9éĆų„ꬌlĦ│÷¼FŻ¼4éĆų„ꬌlĦŽ¹╩¦.▒Ē├„Ż¼▐Dūā┼ÓB£žČ╚║¾Ż¼╝ÜŠ·║═╣┼Š·Ą─Š·╚║ĮYśŗŠ∙│÷¼F├„’@ūā╗»Ż¼Š·╚║ā╚╬ó╔·╬’Ą─╔·╬’┴┐║═ĘNŅÉ│╩¼F┤¾Ę∙Ą─š{š¹Ż¼ęį▀mæ¬ą┬Ą─£žČ╚Śl╝■.
Ī▒ųĖ╩Š▓┐ĘųŻ¼Ž┬═¼);╣┼Š·Š·╚║ųą╝sėą9éĆų„ꬌlĦ│÷¼FŻ¼4éĆų„ꬌlĦŽ¹╩¦.▒Ē├„Ż¼▐Dūā┼ÓB£žČ╚║¾Ż¼╝ÜŠ·║═╣┼Š·Ą─Š·╚║ĮYśŗŠ∙│÷¼F├„’@ūā╗»Ż¼Š·╚║ā╚╬ó╔·╬’Ą─╔·╬’┴┐║═ĘNŅÉ│╩¼F┤¾Ę∙Ą─š{š¹Ż¼ęį▀mæ¬ą┬Ą─£žČ╚Śl╝■.
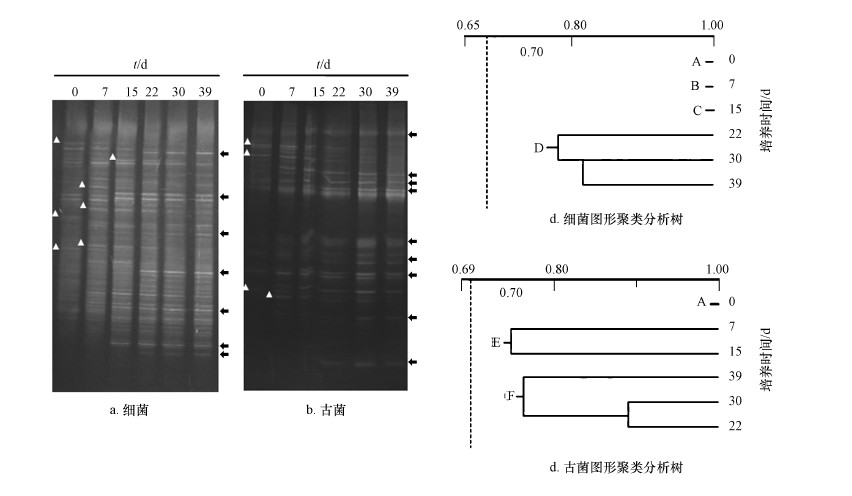
łD 4 DGGE╝╝ągĘų╬÷╝ÜŠ·║═╣┼Š·Ą─Š·╚║ĮYśŗ(ūóŻ║a. ╝ÜŠ·▓╗═¼┼ÓBĢrķgĄ─DGGEłDŽ±;b. ╣┼Š·▓╗═¼┼ÓBĢrķgĄ─DGGEłDŽ±Ż¼Ī░Ī„Ī▒ųĖ╩Šą┬į÷ŚlĦŻ¼Ī░ ![]() Ī▒ųĖ╩ŠŽ¹╩¦ŚlĦ;c. ╝ÜŠ·DGGEłDŽ±Ą─Š█ŅÉĘų╬÷;d. ╣┼Š·DGGEłDŽ±Ą─Š█ŅÉĘų╬÷Ż¼▓╔ė├Quantity One 4.3▄ø╝■UPGMAĘų╬÷.)
Ī▒ųĖ╩ŠŽ¹╩¦ŚlĦ;c. ╝ÜŠ·DGGEłDŽ±Ą─Š█ŅÉĘų╬÷;d. ╣┼Š·DGGEłDŽ±Ą─Š█ŅÉĘų╬÷Ż¼▓╔ė├Quantity One 4.3▄ø╝■UPGMAĘų╬÷.)
ĪĪĪĪ└¹ė├Quantity One▄ø╝■ī”Ę┤æ¬Ų„Š·╚║DGGEłDŲ¼ųąĄ─ŚlĦłDą╬▀MąąĘų╬÷Ż¼╦∙½@öĄō■└¹ė├UPGAMAĘĮĘ©▀MąąŠ█ŅÉĘų╬÷└LųŲ│╔ŽĄĮyśõŻ¼ĮY╣¹ęŖłD 4c.╝ÜŠ·Š·╚║Ą─DGGEłDūV▒╗äØĘų╚ļ4éĆūÕŻ¼0Īó7Īó15 dłDūVĘųäeī┘ė┌AĪóBĪóC 3éĆūÕŻ¼22Īó30Īó39 dłDūV▒╗äØÜw╚ļDūÕŻ¼▒Ē├„į┌▐D╚ļĖ▀£ž┼ÓBĄ─Ū░22 dŻ¼░lĮ═¾wŽĄā╚Ą─Š·╚║ĮYśŗ▓╗öÓš{š¹Ż¼ęį▀mæ¬ą┬£žČ╚ĦüĒĄ─▀xō±ē║┴”.22 d║¾Ż¼Š·╚║ĮYśŗ┌ģė┌ĘĆČ©.╣┼Š·Š·╚║Ą─DGGEłDūV▒╗äØÜw╚ļ3éĆūÕ(łD 4d)Ż¼0 dūVłDäØÜw×ķAūÕŻ¼7 d║═15 dłDūVäØÜw╚ļBūÕŻ¼22Īó30║═39 dłDūVäØÜw╚ļCūÕ.▒Ē├„Ė─ūā£žČ╚┼ÓB║¾Ż¼╣┼Š·Š·╚║ĮYśŗ╝┤«a╔·ūā╗».Š·╚║ĮYśŗĄ─ūā╗»│╠Č╚╚§ė┌╝ÜŠ·Ż¼Ūęūā╗»ų„ę¬╝»ųąį┌┼ÓBĄ─Ū░7 d.┼c╝ÜŠ·ŽÓ╦ŲŻ¼╣┼Š·Š·╚║ĮYśŗūį22 d║¾▀M╚ļĘĆČ©ļAČ╬Ż¼ų▒ų┴┼ÓBĮY╩°Ż¼Š·╚║ĮYśŗø]ėą│÷¼F’@ų°Ą─▓Ņ«É.═©▀^╝ÜŠ·║═╣┼Š·DGGEłDūVūā«ÉĘų╬÷ĮY╣¹Ż¼┐╔░l¼FŻ¼ųą£žģÆ觎¹╗»Š·╚║▐D╚ļĖ▀£ž┼ÓB║¾Ż¼Š·╚║ĮYśŗīóĢ■░l╔·’@ų°Ą─ūā«ÉŻ¼▀@ĘNŠ·╚║ĮYśŗĄ─ūā«ÉīóĢ■│ų└m20 dū¾ėęŻ¼▀@┼cĘ┤款wŽĄšėÜŌ«a┴┐ŽÓ╬Ū║Ž(łD 2d).╝ÜŠ·Ą─ūā▀w│╠Č╚ŽÓī”ė┌╣┼Š·Ė³╝ėäĪ┴ęŻ¼ī¦ų┬▀@ę╗¼FŽ¾Ą─įŁę“┐╔─▄╩Ūė╔ė┌╝ÜŠ·Š·╚║Ą─ČÓśėąį▌^╣┼Š·Š·╚║Ė³×ķžSĖ╗╦∙ę²ŲĄ─.▀@ę╗Ęų╬÷ĮY╣¹┐╔ė╔ßśī”DGGEłDūVĄ─ų„│╔ĘųĘų╬÷Ė³×ķą╬Ž¾Ą─šf├„(łD 5).
ĪĪĪĪłD 5(Fig. 5)
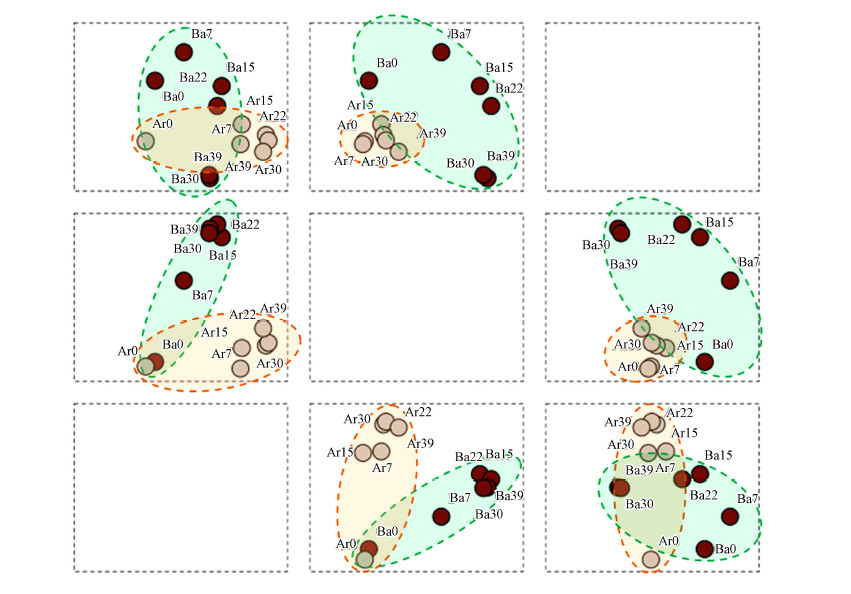
ĪĪĪĪłD 5╝ÜŠ·║═╣┼Š·DEEGŚlĦūā╗»Ą─ų„│╔ĘųĘų╬÷(ūóŻ║łDųąBa×ķ╝ÜŠ·Ż¼Ar×ķ╣┼Š·Ż¼öĄųĄ×ķ┼ÓBĢrķg)
ĪĪĪĪų„│╔ĘųĘų╬÷│Żė├üĒĘų╬÷Š·╚║ĮYśŗūā╗».į┌▒ŠčąŠ┐ųąŻ¼╝ÜŠ·║═╣┼Š·Š·╚║Ą─Ęų╬÷ĮY╣¹▒╗╠ß╚Ī┴╦3éĆų„│╔ĘųŻ¼Ųõ└█ĘeĘĮ▓Ņ×ķ73.2%(ų„│╔Ęų1Īó2Īó3Ęųäe×ķ32.1%Īó28.1%Īó13.0%).łD 5×ķ3éĆų„│╔ĘųĄ─▌d║╔łDŻ¼į¬╦žŅÉą═ŠžĻć.łDųąĮY╣¹’@╩ŠŻ¼╝ÜŠ·Š·╚║║═╣┼Š·╚║į┌łDųą─▄ē“▌^×ķ├„’@Ą─▒╗ģ^Ęų.┼c╣┼Š·Š·╚║ŽÓ▒╚Ż¼╝ÜŠ·Š·╚║į┌▐Dūā┼ÓB£žČ╚║¾Š·╚║ĮYśŗĘĮ▓ŅųĄĖ³×ķĘų╔óŻ¼▒Ē├„╝ÜŠ·Š·╚║Ą─ĮYśŗūā«ÉĖ³×ķÅŖ┴ę.▐D╚ļĖ▀£ž┼ÓB║¾Ż¼2ĮMŠ·╚║Ą─Š·╚║ĮYśŗ┼cįŁ╩╝┼ÓB£žČ╚Š∙«a╔·┴╦├„’@Ą─ūā╗»Ż¼į┌łDųąät▒Ē¼F×ķ0 dĄ─Š·Š·╚║ĮYśŗ║═║¾Ų┌┼ÓBĄ─Š·╚║ĮYśŗŠ∙«a╔·┴╦▌^▀hĄ─ūā«ÉŠÓļx.
ĪĪĪĪ3.4 Ė▀£ž±Z╗»ģÆ觎¹╗»Š·╚║▀^│╠ųąŠ·╚║ČÓśėąįĘų╬÷
ĪĪĪĪģÆ觎¹╗»Š·╚║Ą─16S rRNA╗∙ę“Ų¼Č╬└¹ė├┐╦┬ĪĘĮĘ©▒╗½@╚ĪŻ¼▓óĮ©┴ó┐╦┬Ī╬─ÄņĪó└LųŲŽĄĮy░lė²śõŻ¼½@╚ĪŠ·╚║ā╚Š·╚║ČÓśėąįą┼Žó.
ĪĪĪĪłD 6a’@╩ŠģÆ觎¹╗»Š·╚║ųą╝ÜŠ·Š·╚║Ą─ŽĄĮy░lė²╠žš„.╦∙ėą╝ÜŠ·Š·╚║äØ╚ļ7éĆūÕŻ¼░³└©CaloramatorĪóThermoanaerobacterĪóPeptococcaceaeĪóThermacetogeniumĪóAcetomicrobiumĪóSoehngeniaĪóBacteroides.ŲõųąBa4ūÕų„ę¬×ķThermacetogeniumŠ·Ż¼į┌Š·╚║ųąĄ─▒╚└²ūŅČÓŻ¼▀_ĄĮ33.44%.Ą┌4ūÕųąĄ─ŅÉ╦ŲŠ·ųĻThermacetogenium phaeum strain PB.Ęųļxė┌55 ĪµĖ▀£žĄ─ģÆč§Ę┤æ¬Ų„Ż¼─▄ē“┼c╩╚¤ßÜõĀIBą═«a╝ū═ķŠ·╣▓╔·Č°č§╗»ęę╦ß.Bac5į┌╝ÜŠ·Š·╚║ųąĄ─▒╚└²╝s×ķ20.99%.┤╦ūÕĄ─ęčų¬┤·▒ĒŠ·ųĻ×ķAcetomicrobium faecalŻ¼įōŠ·░l¼Fė┌Ž┬╦«Ą└╬█─ÓŻ¼╦³─▄ē“┤·ųxČÓĘN╝║╠Ū║═╬ņ╠Ū×ķ╚ķ╦ßĪóęę╦ßĪóęę┤╝ĪóCO2Ą╚.┤╦═ŌŻ¼Bac3ĪóBac1ĪóBac7Ą╚ūÕĄ─Š·ųĻį┌ģÆ觎¹╗»Š·╚║ųąš╝ėą▌^┤¾▒╚└²Ż¼Ęųäe×ķ13.5%Īó12.35%Īó12.34%.╦³éāČÓöĄ░l¼Fė┌Ė▀£žģÆ解hŠ│.
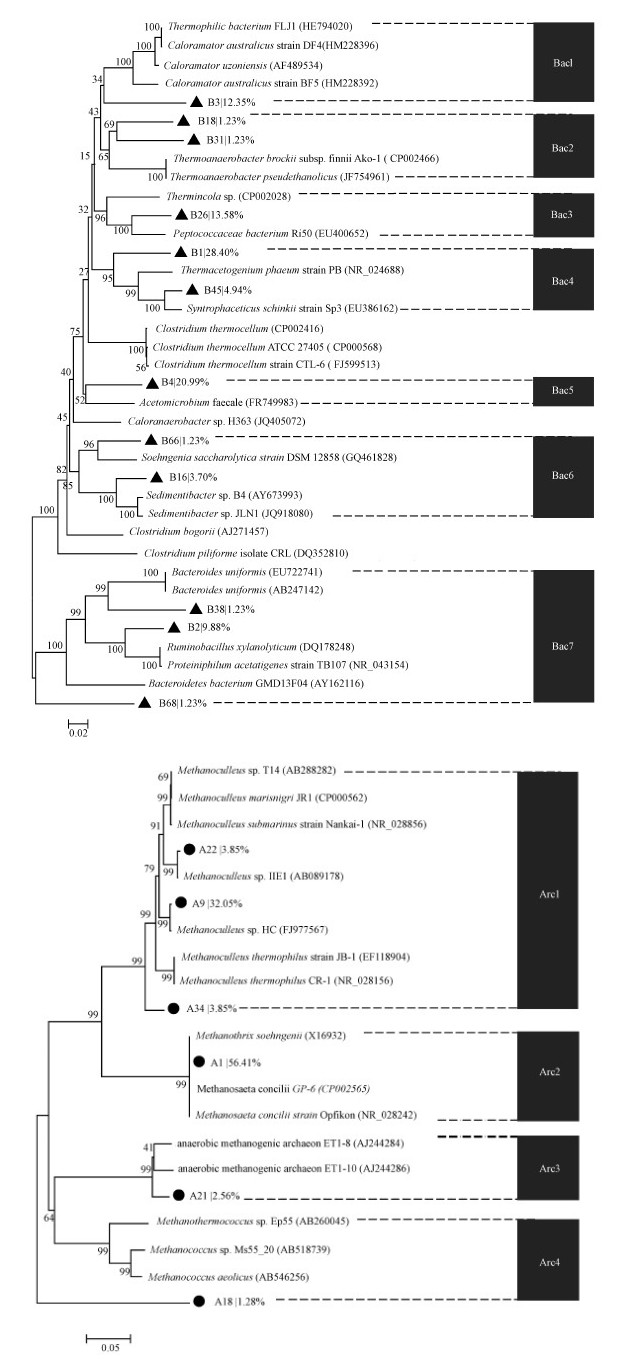
ĪĪĪĪłD 6╝ÜŠ·Īó╣┼Š·Š·╚║16S rDNAą“┴ąĄ─ŽĄĮy░lė²śõĘų╬÷
ĪĪĪĪģÆ觎¹╗»Š·╚║ųąĄ─╣┼Š·Š·╚║▒╗äØĘų×ķ4éĆūÕ(łD 6b)Ż¼ŲõųąArc2×ķ4ūÕųąūŅ×ķžSĖ╗Ą─╣┼Š·Š·╚║Ż¼žSČ╚×ķÖz£y╣┼Š·┐é┴┐Ą─56.4%Ż¼┤·▒ĒŠ·ųĻMethanosaeta concili╩Ūę╗ŅÉ─▄ē“└¹ė├ęę╦ßū„×ķ╬©ę╗─▄į┤Ąū╬’┤·ųx«a╔·Ą╚┴┐Ą─CH4║═CO2.─▄ē“└¹ė├╝ū╦ßĪóęę╦ß║═CO2ū„×ķ╠╝į┤Č°╔·ķL.Arc1į┌╣┼Š·clone╬─Äņųą╦∙š╝Ą─▒╚└²×ķ39.75%Ż¼┤·▒ĒŠ·ųĻ×ķMethanoculleusŻ¼įōŠ·ųĻ─▄ē“└¹ė├H2/CO2ū„×ķĄū╬’Č°╔·ķL▓ó«a╔·╝ū═ķ.ę╗ą®Š·ĘN▀ĆąĶę¬ęę╦ßū„×ķ╔·ķLę“ūė.┤╦═ŌŻ¼▀Ćėąę╗ą®ģÆ觫a╝ū═ķ╣┼Š·╝░Methanococcus(Arc3║═Arc4)▒╗Öz£yĄĮŻ¼╦³éāį┌clone╬─ÄņųąĄ─▒╚└²Ęųäe×ķ2.56%║═1.28%.JonesĄ─蹊┐░l¼FMethanococcus╩Ūę╗ĘN«a╝ū═ķ╣┼Š·Ż¼─▄ē“└¹ė├H2║═CO2╔·│╔╝ū═ķ.Arc3ųąĄ─Į³Šē╣┼Š·×ķChin╦∙ł¾Ą└Ż¼╩Ū═©▀^┐╦┬Ī╬─ÄņĘĮĘ©ė┌╦«╠’═┴╚└ųą░l¼FĄ─Š·ųĻ.
ĪĪĪĪ┐╔ęŖŻ¼▐D╚ļĖ▀£ž┼ÓB║¾Ż¼Š·╚║ųąų„ꬥ─śŗ│╔Š·ųĻ▐DōQ│╔×ķ─▄ē“į┌Ė▀£žŚl╝■Ž┬╔·ķL▓ó┼c╠ŪĪóČÓĘNėąÖC╦ß┤·ųxŽÓĻPĄ─╝ÜŠ·Š·ųĻęį╝░«a╝ū═ķ╣┼Š·Š·ųĻ.š²╩Ūė╔ė┌▀@ą®Š·ųĻĄ─ĘĆČ©┤µį┌Ż¼▓┼─▄ē“ŠS│ųŠ·╚║į┌Ė▀£žŚl╝■Ž┬ĘĆČ©Ž¹╗»ėąÖCÅU╦«╔·│╔šėÜŌ.
ĪĪĪĪ3.5 Ė▀£ž±Z╗»ģÆ觎¹╗»Š·╚║▀^│╠ųąų„ꬊ·ųĻĄ─Č©┴┐Ęų╬÷
ĪĪĪĪ«a╝ū═ķ╣┼Š·Ą─┐é╔·╬’┴┐╝░ę╗ą®Ąõą═Ą─«a╝ū═ķ╣┼Š·Ą─╔·╬’┴┐┐╔ęį└¹ė├¤╔╣ŌČ©┴┐PCRĘĮĘ©▀MąąČ©┴┐Ęų╬÷.ĮY╣¹’@╩ŠŻ¼ģÆ觎¹╗»Š·╚║Å─37 Īµ▐DęŲų┴50 ĪµŻ¼«a╝ū═ķ╣┼Š·Ą─┐é╔·╬’┴┐£p╔┘(łD 7c)Ż¼▓óį┌22 d║¾▀M╚ļŽÓī”ĘĆČ©ĀŅæBŻ¼║¼┴┐╝s×ķ7.6Ī┴106 ┐ĮžÉ/g╗Ņąį╬█─ÓŻ¼ŽÓ▒╚ė┌ųą£ž┼ÓB£p╔┘42.2%.└¹ė├╠ž«Éąį¤╔╣Ō╠ĮßśŻ¼Öz£y┴╦MMBŻ¼MCCŻ¼MST 3ŅÉ«a╝ū═ķ╣┼Š·Ż¼░l¼FŻ¼MMB╩Ūę╗ŅÉį┌37 Īµ║═50 ĪµŠ∙─▄Öz£yĄĮŻ¼▓ó┤¾┴┐┤µį┌Ą─«a╝ū═ķ╣┼Š·ŅÉ╚║Ż¼▀@┼cČŁ╗█ė°Ą╚(2014)Ą─蹊┐ĮY╣¹ŽÓę╗ų┬.MCC║═MSTĄ─╔·╬’┴┐į┌╚║¾wųą
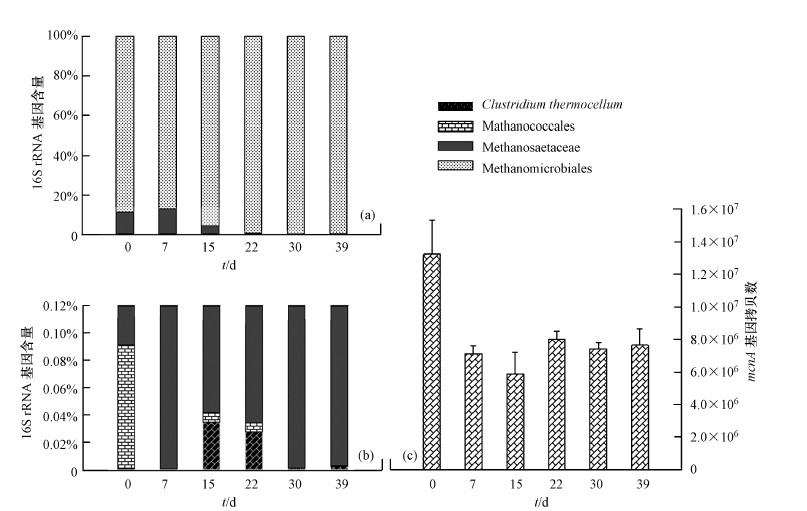
ĪĪĪĪłD 7ģÆ觎¹╗»Š·╚║ā╚«a╝ū═ķ╣┼Š·Ą─╔·╬’┴┐(ūóŻ║(a).Ė„ŅÉŠ·ųĻĄ─16S rRNA╗∙ę“┐ĮžÉöĄš╝▒╗Öz£y┐銷ųĻ┐ĮžÉöĄĄ─▒╚└²;(b)Ż¼(a)Ę┼┤¾ĄĮ0.12%(Y▌S)│▀Č╚Ą─łDŽ±;(c).«a╝ū═ķ╣┼Š·mcrA╗∙ę“Ą─┐ĮžÉöĄ)
ĪĪĪĪ▌^╔┘╝sš╝Öz£y4ĘNŠ·ųĻ┐é┴┐Ą─11%Ż¼▓óŪęį┌▐D╚ļĖ▀£ž┼ÓB║¾Ż¼Ųõ╔·╬’┴┐Ą─╝▒äĪ£p╔┘Ż¼MCC╔§ų┴į┌Š·╚║ĘĆČ©║¾Ž¹╩¦.▒Ē├„▀@2ĘN«a╝ū═ķ╣┼Š·▓ó▓╗▀mę╦ė┌50 ĪµŚl╝■Ž┬╔·ķL.Clostridium thermocellum╩Ūę╗ųĻć└Ė±ģÆ觥─└wŠS╦žĘųĮŌŠ·ųĻŻ¼ÅVĘ║┤µį┌ė┌║¼ėą└wŠS╦ž╗∙┘|Ą─ģÆč§ĮĄĮŌŁhŠ│.C. thermocellum ╩Ūę╗ųĻ─▄ē“į┌Ė▀£ž┴╝║├╔·ķLĄ─Š·ųĻŻ¼į┌▒Š╬─ųąŻ¼▐D╚ļĖ▀£ž║¾╔·╬’┴┐Ę┤Č°ąĪė┌│Ż£ž┼ÓBŻ¼═Ų£y×ķģÆ觎¹╗»¾wŽĄęį▓╗║¼└wŠS╦ž╠Ū├█ÅU╦«×ķų„Ż¼ę“╚▒Ę”▀mę╦Ą─┤·ųxĄū╬’Ż¼ļSų°ģÆ觎¹╗»¾wŽĄ▀\ąąĢrķgĄ─čėķLŻ¼╔·╬’┴┐ųØu╦ź═╦.
ĪĪĪĪ4 ėæšō
ĪĪĪĪį┌ģÆ觎¹╗»▀^│╠Ż¼£žČ╚═©│Ż╩Ūė░Ēæ╬ó╔·╬’Š·╚║Ą─’@ų°ę“ūėŻ¼ė░Ēæ╬ó╔·╬’Ą─╔·ķL║═╔·┤µ.▒ŠčąŠ┐īó37 ĪµĄ─ģÆ觎¹╗»Š·╚║ų▒Įė╠ߣžų┴50 Īµ±Z╗»┼ÓBŻ¼į┌£žČ╚║═Ąū╬’Ą─▀xō±ē║┴”Ž┬Ż¼įŁ╩╝Š·╚║ĮYśŗĮø▀^äĪ┴ęĄ─ūā╗»š{š¹║¾ųØu▀mæ¬ą┬Ą─£žČ╚Śl╝■Ż¼▓óŠ▀ėą«a╝ū═ķĄ─╣”─▄.═©│Żį┌ģÆ觎¹╗»Š·╚║±Z╗»ųąŻ¼ČÓ▓╔ė├ųą£žŠ·╚║╠▌Č╚╔²£žĄ─ĘĮĘ©½@Ą├Ė▀£žģÆ觎¹╗»Š·╚║Ż¼╚ńÜWĻ¢Č■├„Ą╚Ą╚▓╔ė├ą“╠▌Č╚╔²£žĘĮĘ©īóųą£ž╗Ņąį╬█─Ó(35 Īµ)±Z╗»×ķĖ▀£ž╠Ä└Ē¤ß╦«ĮŌ╬█─ÓĄ─Š·╚║.└Ņ│¼éźĄ╚ęį╩│╠├▓═ÅN└¼╗°×ķįŁ┴ŽŻ¼▓╔ė├ųą£žšėę║ū„×ķŠ·ĘNĮøŠÅ┬²╔²£ž│╔╣”åóäė┴╦Ė▀£ž╠Ä└Ē▓═ÅN└¼╗°Ą─ģÆ觎¹╗»čbų├.į┌▒ŠčąŠ┐ųą═©▀^ę╗▓Įų▒Įė╠ߣžĄ─ĘĮĘ©ę▓┐╔½@Ą├Ė▀£žģÆ觎¹╗»Ė▀ØŌČ╚ėąÖCÅU╦«Š·╚║Ż¼▀@┼cAhringĪóTian║═RaskinĄ─ė^³cŽÓę╗ų┬.
ĪĪĪĪę╗ą®ųą£žģÆ觎¹╗»Š·╚║ųą│Ż┤µį┌▀mę╦ė┌Ė▀£ž╔·ķLĄ─╣”─▄Š·ųĻŻ¼▀@▓┐ĘųŠ·ųĻ▓ó▓╗Ģ■į┌ųą£žŚl╝■Ž┬Ą─ķLŲ┌┼ÓBČ°üG╩¦.¤ošō▒ŠčąŠ┐ųąīóĘųĮŌŪ▌ą¾╝S▒ŃĄ─ųą£žģÆ觎¹╗»Š·╚║ų▒Įė▐D╚ļĖ▀£ž┼ÓBŻ¼½@Ą├Ė▀£žģÆ觎¹╗»Š·╚║Ż¼▀Ć╩Ū└¹ė├╠▌Č╚╔²£žĘ©½@Ą├Ė▀£žģÆ觎¹╗»Š·╚║Ż¼ŲõĮY╣¹Š∙─▄╩╣┤µį┌ė┌ųą£žŠ·╚║ųąĄ─Ė▀£ž╣”─▄Š·ųĻ╗Ņ╗»▓óĖ╗╝»×ķā×ä▌Š·╚║Č°░lō]╣”─▄.Ųõģ^äeį┌ė┌Š·╚║į┌£žČ╚Ė─ūā▀^│╠ųą╦∙├µ┼RĄ─▀xō±ē║┴”┤¾ąĪ.’@╚╗Ż¼ų▒Įė╠ß╔²£žČ╚Ą─ĘĮĘ©į÷╝ė┴╦▀@ĘN▀xō±ē║┴”.
ĪĪĪĪį┌Š·╚║ĮYśŗūā▀w▀^│╠ųąŻ¼╝ÜŠ·Ą─ūā╗»│╠Č╚▌^╣┼Š·ÅŖ┴ę.GobemaĄ╚蹊┐šJ×ķŻ¼╝ÜŠ·Š▀ėąĘŪ│ŻĖ▀Ą─ČÓśėąįŻ¼į┌│Ż£žģÆč§Ę┤æ¬Ų„ā╚Ż¼ŲõĘNŅÉžSĖ╗│╠Č╚▀hĖ▀ė┌╣┼Š·.▒ŠčąŠ┐ųąŻ¼«ö┼ÓB£žČ╚╠ßĖ▀ęį║¾Ż¼┤¾┴┐DGGEŚlĦ╗“Ž¹╩¦Īó╗“ą┬į÷Ż¼▒Ē├„Š·╚║ā╚ę╗ą®¤oĘ©▀mæ¬ą┬┼ÓB£žČ╚Š·ųĻĄ─╔·╬’┴┐¾E£pŻ¼Č°▀mæ¬Ė▀£ž╔·ķL╝ÜŠ·Š·ųĻĄ─╔·╬’┴┐į÷╝ė▓óš╝ō■┴╦╗Ņ▄SĄ─ų„ī¦Ąž╬╗.┼c╝ÜŠ·Š·╚║ŽÓ▒╚▌^Ż¼╣┼Š·Š·╚║Ą─ČÓśėąįūā«É▌^×ķŲĮŠÅŻ¼▀@│²┴╦┼c╣┼Š·ĘNŅÉ▌^╔┘(łD 3Īó5Īó6)ėąĻPŻ¼┐╔─▄ę▓║═╣┼Š·╔·ķLų▄Ų┌ķLĪó╔·ķLŠÅ┬²ėąĻP.╝ÜŠ·ę“Ųõ╔·ķL╦┘Č╚┐ņŻ¼Š·╚║ĮYśŗĖ³ą┬ų▄Ų┌Č╠Ż¼▒Ē¼F×ķ╝ÜŠ·Š·╚║ĮYśŗūā«ÉäĪ┴ę.Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.bnynw.comĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
ĪĪĪĪ5 ĮYšō
ĪĪĪĪ1) 37 Īµ┼ÓBĄ─ųą£žģÆ觎¹╗»«a╝ū═ķŠ·╚║ų▒Įė▐D╚ļ50 ĪµĖ▀£ž±Z╗»╠Ä└ĒĖ▀ØŌČ╚╠Ū├█ėąÖCÅU╦«Ż¼ģÆ觎¹╗»▀^│╠─▄ē“┐ņ╦┘åóäė▓ó╔·│╔╝ū═ķŻ¼▓óį┌22 d║¾ą╬│╔ĘĆČ©Ą─Ė▀£žģÆ觎¹╗»«a╝ū═ķŠ·╚║Ż¼ŲĮŠ∙╝ū═ķ╔·│╔ą¦┬╩×ķ162.5 mLĪżg-1.ĘĆČ©«aÜŌŲ┌ķgęę╦ß║═▒¹╦ߥ─┘|┴┐ØŌČ╚Ęųäe×ķ25.3║═145.3 mgĪżL-1.
ĪĪĪĪ2) Š·╚║ĮYśŗ╝░ČÓśėąįį┌Ė▀£ž±Z╗»▀^│╠ųą░l╔·äĪ┴ęūā╗»Ż¼╝ÜŠ·ÅŖė┌╣┼Š·Ż¼▓óųØuĘĆČ©│╔×ķęį╝ÜŠ·(ThermacetogeniumŻ¼Acetomicrobium)ęį╝░«a╝ū═ķ╣┼Š·(MethanosaetaŻ¼Methanoculleus)×ķų„ę¬ā×ä▌Š·╚║Ą─Ė▀£žģÆ觎¹╗»Š·╚║.
ĪĪĪĪ3) ╦∙½@Ė▀£žģÆ觎¹╗»«a╝ū═ķŠ·╚║Ż¼Ųõ«a╝ū═ķ╣┼Š·Ą─┐é╔·╬’┴┐├„’@Ž┬ĮĄŻ¼┤¾╝s×ķ7.6Ī┴106┐ĮžÉ/g╗Ņąį╬█─Ó.

