ņoų├/║├č§/╚▒觹“┼·╩ĮĘ┤æ¬Ų„(SBR)├ōĄ¬│²┴ū╝╝ąg
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2016-7-9 8:58:44
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ1 ę²čį
ĪĪĪĪĄõą═Ą─įOŪ░ų├Ę┤Ž§╗»Č╬Ą─╔·╬’├ōĄ¬│²┴ū╣ż╦ćėąģÆč§/╚▒č§/║├č§╣ż╦ć(A2/O╣ż╦ć)ĪóUniversity of Cape Town╣ż╦ć(UTC╣ż╦ć)╝░╔·╬’╗»īW├ōĄ¬│²┴ū╣ż╦ć(BCFS╣ż╦ć).Ę┤Ž§╗»Č╬Ū░ų├Ą─ā×ä▌╩ŪģÆč§║Ž│╔Ą─ā╚Š█╬’Š█”┬┴u╗∙ų¼ĘŠ╦ß(PHA)Ą╚┐╔ų▒Įė▀M╚ļ╚▒č§Č╬“īäėĘ┤Ž§╗»Č°╚ĪĄ├▌^║├Ą─├ōĄ¬ą¦╣¹Ż¼Ą½Ū░ų├Ę┤Ž§╗»Č╬ėąŲõ╣╠ėąĄ─╚▒Ž▌.Ė∙ō■╔·╬’├ōĄ¬└ĒšōŻ¼Ž§╗»Č╬(║├č§Č╬)ā╚░▒č§╗»Š·(AOBs)īó░▒¹}č§╗»×ķü厧╦ß¹}║¾Ż¼ü厧╦ß¹}č§╗»Š·(NOBs)īóü厧╦ß¹}č§╗»×ķŽ§╦ß¹};Ę┤Ž§╗»Č╬(╚▒č§Č╬)ā╚Ę┤Ž§╗»Š·īóŽ§╦ß¹}▀ĆįŁ×ķü厧╦ß¹}Ż¼▓ó▀Mę╗▓Į▀ĆįŁ×ķĄ¬ÜŌ(N2)(Zhou et al., 2011).ė╔ė┌║├č§Č╬į┌╚▒č§Č╬║¾Ż¼×ķīŹ¼FĘ┤Ž§╗»Ż¼ę“Č°▒žĒÜīó╗ņ║Žę║Å─║├č§Č╬╗ž┴„ų┴╚▒č§Č╬.╗ņ║Žę║╗ž┴„Ģ■ŽĪßī▀M╦«ėąÖC┘|ØŌČ╚;č§╗»æBĄ¬(NO-x)Ą─╚ź│²ę▓╩▄ųŲė┌╗ņ║Žę║Ą─╗ž┴„╦┘┬╩Ż¼Ūę═Ļ╚½├ōĄ¬▓╗┐╔─▄īŹ¼F;╗ņ║Žę║╗ž┴„▀ĆĢ■į÷╝ė─▄┴┐Ž¹║─║═╣ż╦ćÅ═ļsČ╚.
ĪĪĪĪ╚▒č§Č╬ų├ė┌║├č§Č╬ų«║¾Ą─║¾ų├╚▒č§Ę┤Ž§╗»ĘĮ╩ĮŻ¼ę“╩Ī╚ź┴╦╗ņ║Žę║╗ž┴„Č°║å╗»┴╦╣ż╦ć┴„│╠Ż¼Ūę─▄īŹ¼F▌^║├Ą─├ōĄ¬│²┴ūą¦╣¹Č°Ą├ĄĮ┴╦ÅVĘ║Ą─蹊┐.┼cŪ░ų├Ę┤Ž§╗»ŽÓ▒╚Ż¼═Ō╠╝į┤ęčį┌ģÆč§Č╬╗“║├č§Č╬Ž¹║─Ż¼║¾ų├╚▒č§Č╬Ę┤Ž§╗»Š·ęįā╚╠╝į┤(╠ŪįŁ╗“PHA)×ķļŖūė╣®¾wŻ¼ęįNO-x×ķļŖūė╩▄¾w“īäėĘ┤Ž§╗».Coats Ą╚(2011)║═WinklerĄ╚(2011)蹊┐┴╦║¾ų├╚▒觹“┼·╩ĮĘ┤æ¬Ų„(SBR)╣ż╦ćŻ¼BracklowĄ╚(2010)║═VocksĄ╚(2005)蹊┐┴╦▀B└m┴„║¾ų├╚▒č§─ż╔·╬’Ę┤æ¬Ų„(MBR)╣ż╦ćŻ¼Š∙╚ĪĄ├┴╦┴╝║├Ą─├ōĄ¬│²┴ūą¦╣¹.▀@ą®čąŠ┐▒Ē├„Ż¼║¾ų├╚▒č§Č╬ļm╬┤═Ō╝ė╠╝į┤Ż¼Ą½╬ó╔·╬’┐╔└¹ė├░¹ā╚╠ŪįŁ╗“PHA“īäėĘ┤Ž§╗»├ōĄ¬.┤╦═ŌŻ¼XuĄ╚(2011)į┌║¾ų├╚▒č§Ę┤Ž§╗»Ą─╗∙ĄA╔Žīó▓┐ĘųģÆč§Č╬╗ņ║Žę║Ęų┼õ▀M╚▒č§Č╬īŹ¼FĘ┤Ž§╗»│²┴ūŻ¼▓óį┌║├č§Č╬īŹ¼F┴╦═¼▓ĮŽ§╗»-Ę┤Ž§╗».▀@ĘNĖ─▀Mļm╚╗īŹ¼F┴╦Ę┤Ž§╗»│²┴ūŻ¼Ą½ėųį÷╝ė┴╦╣ż╦ćÅ═ļs│╠Č╚.║¾ų├Ę┤Ž§╗»Ą─ģÆč§/║├č§/╚▒č§SBR╣ż╦ćĮŌøQ┴╦╗ņ║Žę║╗ž┴„Ą─å¢Ņ}Ż¼Ą½įō╣ż╦ć╩Ūʱėą▀Mę╗▓ĮĖ─▀MĄ─┐šķg?
ĪĪĪĪĖ∙ō■é„Įy╔·╬’ÅŖ╗»│²┴ū(EBPR)└ĒšōŻ¼EBPR═©▀^ģÆč§/║├č§╗“ģÆč§/╚▒č§Į╗╠µ▀\ąąīŹ¼F.▀@ĘNŚl╝■×ķŠ█┴ūŠ·(PAOs)┤·ųx╔·ķL╠ß╣®▀xō±ąįā×ä▌Ż¼╩╣ų«─▄ģÆč§╬³╩šō]░ląįų¼ĘŠ╦ß(VFAs)║Ž│╔×ķPHAŻ¼▓ó║├č§╬³╩š┴ū╦ß¹}(Coats et al., 2011).PAOsģÆč§╬³╩šVFAsĄ──▄┴┐üĒį┤ė┌Š█┴ūĮĄĮŌ║═╠ŪįŁĘųĮŌŻ¼Č°╠ŪįŁĘųĮŌ×ķPHA║Ž│╔╠ß╣®▀ĆįŁ┴”(Smolders et al., 1994).į┌║├č§╗“╚▒觌l╝■Ž┬Ż¼PAOs═©▀^╚²¶╚╦ß裣h(TCA)×ķūį╔Ē╔·ķLĪó╠ŪįŁā”┤µĪó┴ū╦ß¹}öz╚Ī║═Š█┴ū║Ž│╔╠ß╣®─▄┴┐(Smolders et al., 1995).EBPRŽĄĮyųąę▓┤µį┌Š█╠ŪŠ·(GAOs)Ż¼▀@ĘN╬ó╔·╬’│²▓╗─▄ģÆč§ßī┴ū║═║├č§öz┴ū═ŌŻ¼Ųõ╦¹┤·ųxĘĮ╩Į┼cPAOsŽÓ╦ŲŻ¼╣╩─▄┼cPAOsą╬│╔ĖéĀÄĻPŽĄŻ¼ė░Ēæ│²┴ūą¦╣¹.ę“┤╦Ż¼×ķ▀_ĄĮŠ█┴ūŠ·ßī┴ūĄ──┐Ą─Ż¼╗Ņąį╬█─ÓąĶĮøģÆč§öć░Ķęį│õĘųĮėė|╬█╦«ųąVFAs.Š█┴ūŠ·─▄═©▀^Š█┴ūĘųĮŌ╣®─▄╬³╩šVFAsŻ¼Ą½╣Pš▀蹊┐ģÆč§/║├č§/╚▒č§SBR╣ż╦ćĢr░l¼FŻ¼▀M╦«║¾╬┤ģÆč§öć░ĶČ°ņoų├1 h║¾ų▒ĮėŲžÜŌŻ¼ņoų├Ų┌ŽĄĮyųą╚į┐╔▒O£yĄĮ┴ū╦ß¹}Ą─┤¾┴┐ßīĘ┼Ż¼ŪęŲžÜŌķ_╩╝║¾┴ū╦ß¹}╚į─▄▒╗┐ņ╦┘▀^┴┐╬³╩š.═¼ĢrŻ¼║¾ų├╚▒č§Č╬īŹ¼F┴╦Ę┤Ž§╗»├ōĄ¬Ż¼Å─Č°▀_ĄĮ┴╦├ōĄ¬│²┴ūĄ──┐Ą─.į┌╬┤öć░ĶČ°ų▒Įėņoų├ļAČ╬Ż¼ļm╬┤┼c╬█╦«│õĘųĮėė|ŽĄĮyŻ¼╗Ņąį╬█─Ó×ķ╩▓├┤─▄Ž±é„ĮyģÆč§Č╬─Ūśė┤¾┴┐ßī┴ū?įōļAČ╬Š█┴ūŠ·ßī┴ū┼cé„ĮyģÆč§ßī┴ūėą╩▓├┤«É═¼ų«╠Ä?ė╔ė┌▓ó╬┤öć░ĶŻ¼ņoų├Č╬Š█┴ūŠ·┤·ųxĘĮ╩ĮĢ■▓╗Ģ■┼cé„ĮyģÆč§Č╬Š█┴ūŠ·┤·ųxĘĮ╩Įėą╦∙▓╗═¼?ņoų├Č╬Ą─įOų├ī”║├č§öz┴ū╝░╚▒č§Ę┤Ž§╗»├ōĄ¬ė░Ēæėą──ą®?Ę┤Ž§╗»├ōĄ¬╩Ū▓╗╩Ūę▓═©▀^ā╚Š█╬’“īäėŻ¼╩Ū═©▀^╠ŪįŁ▀Ć╩ŪPHA╗“ā╔š▀Š∙ėą?▀@ą®å¢Ņ}Č╝ųĄĄ├╔Ņ╚ļ蹊┐.═¼ĢrŻ¼╚¶ņoų├/║├č§/╚▒č§SBR┐╔▀_ĄĮ┼cģÆč§/║├č§/╚▒č§SBRŽÓ«ö╔§ų┴Ė³║├Ą─├ōĄ¬│²┴ūą¦╣¹Ż¼ęįņoų├Č╬┤·╠µģÆč§Č╬▓ó╩Īģsöć░ĶŻ¼īó║å╗»╣ż╦ć▓ó╣Ø╩Ī─▄┴┐Ż¼▀@ę▓╩Ūį┌║¾ų├╚▒č§╣ż╦ć╔ŽĄ─▀Mę╗▓Įā×╗»╠Į╦„.▒Ŗ╦∙ų▄ų¬Ż¼Ą¬┴ū┤¾┴┐┼┼Ę┼Ģ■ę²Ų╦«¾wĖ╗ĀIB╗»Ż¼ī¦ų┬╦«┘|É║╗»Ż¼Č°é„Įy╔·╬’├ōĄ¬│²┴ū╣ż╦ć▀\ąą│╔▒ŠĖ▀.ę“┤╦Ż¼▀@ĘN╠Į╦„ī”ė┌║å╗»╣ż╦ćĪó╣Ø╝s│╔▒ŠĪóĘ└ų╬╦«¾wĖ╗ĀIB╗»Š▀ėąųžę¬Ą─└Ēšō║═īŹļHęŌ┴xŻ¼ųĄĄ├Ė³╔Ņ╚ļĄžčąŠ┐Ęų╬÷.
ĪĪĪĪę“┤╦Ż¼▒Š╬─ęįęę╦ßŌc×ķå╬ę╗╠╝į┤Ż¼┐╝▓ņņoų├/║├č§/╚▒č§SBRķLŲ┌▀\ąąųą├ōĄ¬│²┴ūąį─▄Ż¼▓óįOų├ģÆč§/║├č§/╚▒č§SBRęį▀Mąąī”▒╚.ė╔ė┌▒ŠčąŠ┐ęįņoų├Č╬╠µ┤·é„ĮyģÆč§Č╬Ż¼ę“Č°╩ūŽ╚▒žĒÜ╠ĮŠ┐ņoų├Č╬Š█┴ūŠ·┤·ųxÖC└ĒŻ¼ęįšō╩÷ņoų├Č╬┼cé„ĮyģÆč§Č╬Ą─«É═¼³c╝░ņoų├Č╬į┌ņoų├/║├č§/╚▒č§SBRĄ─Ąž╬╗ū„ė├.Ųõ┤╬┐╝▓ņā╔ŽĄĮyßī┴ūĪó║├č§öz┴ūĪó═¼▓ĮŽ§╗»-Ę┤Ž§╗»╝░╚▒č§Ę┤Ž§╗»«É═¼³c.╚╗║¾Ęų╬÷ņoų├Č╬╠µ┤·ģÆč§Č╬ī”ŽĄĮyöz┴ūĪóŽ§╗»╝░Ę┤Ž§╗»ėą──ą®ė░ĒæŻ¼ęį╔Ņ╚ļ蹊┐ņoų├/║├č§/╚▒č§SBRĄ─├ōĄ¬│²┴ūąį─▄.ūŅ║¾Ż¼Ė∙ō■├ōĄ¬│²┴ūą¦╣¹Ż¼ī”ņoų├/║├č§/╚▒č§SBR┐╔ʱ╠µ┤·ģÆč§/║├č§/╚▒č§SBRū„│§▓Į┼ąöÓŻ¼ęįŲ┌×ķ║¾ų├╚▒č§╣ż╦ćĄ─čą░l┼cæ¬ė├╠ß╣®ą┬╦╝┬Ę.
ĪĪĪĪ2 ▓─┴Ž┼cĘĮĘ©
ĪĪĪĪ2.1 įć“×čbų├┼c▀\ąąĘĮĘ©
ĪĪĪĪįć“×į┌2éĆŽÓ═¼Ą─SBRųą▀MąąŻ¼Ę┤æ¬Ų„ėąą¦¾wĘe×ķ1.8 L.ĮėĘNĄ─╗Ņąį╬█─Ó╚ĪūįķL╔│╩ąĄ┌ę╗╬█╦«╠Ä└ĒÅSŻ¼Ų╩╝╗Ņąį╬█─ÓØŌČ╚╝s×ķ4000 mg Īż L-1.R1▀\ąąĘĮ╩Į╚ńŽ┬Ż║▀M╦«Ī·ņoų├(1 h)Ī·ŲžÜŌ(2.5 h)Ī·╚▒č§öć░Ķ(3 h)Ī·│┴ĄĒ│÷╦«(0.5 h)Ī·ķeų├(1 h);R2▀\ąąĘĮ╩Į╚ńŽ┬Ż║▀M╦«Ī·öć░Ķ(1 h)Ī·ŲžÜŌ(2.5 h)Ī·╚▒č§öć░Ķ(3 h)Ī·│┴ĄĒ│÷╦«(0.5 h)Ī·ķeų├(1 h).║├č§▓╔ė├╣─’LŲžÜŌŻ¼ŲžÜŌ┴┐┐žųŲ×ķ1.5 L Īż min-1.╦«┴”═Ż┴¶Ģrķg(HRT)×ķ12 h.┐žųŲ╬█─Ó═Ż┴¶Ģrķg×ķ20 dŻ¼š¹éĆ▀^│╠ųą▓╗┐žųŲpH.
ĪĪĪĪ2.2 ╬█╦«╦«┘|
ĪĪĪĪā╔Ę┤æ¬Ų„▀M╦«▓╔ė├║Ž│╔ÅU╦«Ż¼▀M╦«│╔Ęų╝░ØŌČ╚ę╗ų┬.ęę╦ßŌc(10.88 mmol Īż L-1Ż¼ęįCėŗŻ¼└ĒšōCODųĄ×ķ350 mg Īż L-1)×ķå╬ę╗╠╝į┤Ż¼░▒Ą¬▓╔ė├┬╚╗»õ@(40 mg Īż L-1Ż¼ęįNH+4-Nėŗ)Ż¼╚▄ĮŌąįš²┴ū╦ß¹}(SOP)▓╔ė├┴ū╦ßČ■ÜõŌø(12 mg Īż L-1Ż¼ęįPO3-4-Pėŗ).┤╦═ŌŻ¼CaCl2╝░MgSO4ØŌČ╚Įį×ķ5 mg Īż L-1Ż¼╬ó┴┐į¬╦ž╚▄ę║×ķ0.5 mLŻ¼ŲõĮMĘų╝░ØŌČ╚ęŖ╬─½I(Wang et al., 2008).
ĪĪĪĪ2.3 Ęų╬÷ĘĮĘ©
ĪĪĪĪPHA£yČ©▓╔ė├ÜŌŽÓ╔½ūVĘ©(Wang et al., 2008);NO-2-N£yČ©▓╔ė├N-(1-▌┴╗∙)-ęęČ■░ĘĘų╣Ō╣ŌČ╚Ę©Ż¼SOP£yČ©▓╔ė├ŃfõR┐╣Ęų╣Ō╣ŌČ╚Ę©Ż¼NO-3-N£yČ©▓╔ė├ūŽ═ŌĘų╣Ō╣ŌČ╚Ę©Ż¼╗ņ║Žę║ō]░ląįæęĖĪ╣╠¾wØŌČ╚(MLVSS)£yČ©▓╔ė├ųž┴┐Ę©Ż¼NH+4-N£yČ©▓╔ė├╝{╩Žįćä®Ęų╣Ō╣ŌČ╚Ę©Ż¼Š▀¾wĘĮĘ©ģóšš╬─½I(ć°╝ęŁh▒Ż┐éŠųĪČ╦«║═ÅU╦«▒O£yĘų╬÷ĘĮĘ©ĪĘŠÄ╬»Ģ■Ż¼2002);╬█─Óųą┐é┴ū(TP)▓╔ė├NaOH╚█╚┌ŃfõR┐╣Ęų╣Ō╣ŌČ╚Ę©£yČ©(Wang et al., 2009)Ż¼╠ŪįŁ(Glycogen)▓╔ė├▒ĮĘė-┴“╦ßĘ©£yČ©(Wang et al., 2009)Ż¼ÅU╦«┐éėąÖC╠╝ØŌČ╚(TOC)▓╔ė├TOC£yČ©āx(ShimadzuTOC-500Ż¼╚š▒Š)£yČ©.
ĪĪĪĪ3 ĮY╣¹┼cĘų╬÷
ĪĪĪĪ3.1 ņoų├Č╬╬ó╔·╬’┤·ųxĘų╬÷
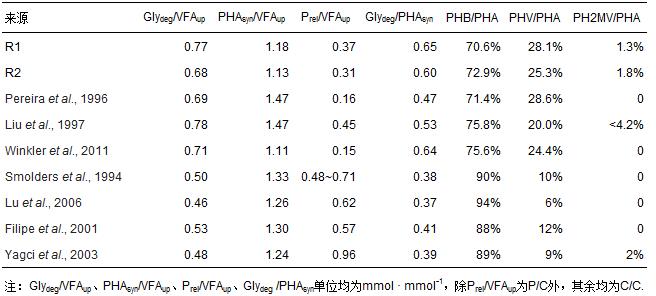
ĪĪĪĪė╔ė┌ņoų├Č╬┼cé„ĮyģÆč§Č╬ėą╦∙▓╗═¼Ż¼Č°▀@ĘN▓Ņ«É╩Ūʱė░Ēæ╬ó╔·╬’┤·ųxŻ¼▀MČ°ė░Ēæ╔·╬’ÅŖ╗»│²┴ūą¦╣¹ųĄĄ├╠Įėæ.▒Š╬─īóÅ─ßī┴ūĪóVFAöz╚ĪĪó╠ŪįŁĘųĮŌ╝░PHA║Ž│╔Ą╚ĘĮ├µ▀MąąĘų╬÷.▀\ąąĘĆČ©ļAČ╬Ż¼R1║═R2Ąõą═ų▄Ų┌ā╚SOPĪóTOCĪó╠ŪįŁ║═PHA(Š█”┬┴u╗∙ČĪ╦ß(PHB)+Š█”┬┴u╗∙╬ņ╦ß(PHV)+Š█Č■╝ū╗∙╚²┴u╗∙╬ņ╦ß(PH2MV))ę╗éĆų▄Ų┌ā╚Ą─ūā╗»Ūķør╚ńłD 1╦∙╩Š.

ĪĪĪĪłD 1 R1(a)║═R2(b)Ąõą═ų▄Ų┌ā╚SOPĪó╠ŪįŁ║═PHAĄ─ūā╗»
ĪĪĪĪR2ģÆč§Č╬čĖ╦┘ßī┴ūŻ¼ßī┴ū┴┐×ķ74.34 mg Īż L-1Ż¼ßī┴ū╦┘┬╩×ķ24.11 mg Īż g-1 Īż h-1(ęį├┐g VSSßīĘ┼Ą─P(mg)ėŗ).R1ņoų├Č╬ßī┴ū┴┐ŽÓī”▌^╔┘Ż¼Ą½╚įėą38.67 mg Īż L-1Ż¼ßī┴ū╦┘┬╩×ķ12.86 mg Īż g-1 Īż h-1. Montil Ą╚(2005)║═MamaisĄ╚(1992)šJ×ķé„ĮyEBPR╣ż╦ćģÆč§Č╬┴ū╦ß¹}ßīĘ┼╦┘┬╩Ęųäe×ķ5~30 mg Īż g-1 Īż h-1║═7~20 mg Īż g-1 Īż h-1. ļm╚╗R1ßī┴ū┴┐āH×ķR2Ą─1/2Ż¼Ą½┼cé„ĮyģÆč§Č╬ßī┴ū╦┘┬╩ŽÓ▒╚Ż¼R1ņoų├Č╬ßī┴ū╦┘┬╩į┌Ųõūā╗»ĘČć·ā╚ŪęŲ½Ė▀.▀@▒Ē├„▀M╦«║¾ļm╬┤öć░Ķ╗Ņąį╬█─ÓŻ¼Ą½R1ųąŠ█┴ūŠ·╚į┐╔▌^┐ņĘųĮŌŠ█┴ūęįčĖ╦┘╬³╩š╦«ųąVFAŻ¼╣╩Ųõņoų├ßī┴ū╦┘┬╩╚į▌^Ė▀.┐╔ęŖŻ¼R1ŽĄĮyā╚ļm¤oé„ĮyģÆč§Č╬Ż¼Š█┴ūŠ·╚į─▄ā×ä▌╔·ķL.┼cR2ģÆč§Č╬ę╗ų┬Ż¼R1ņoų├Č╬ę▓Ģ■░l╔·╠ŪįŁĘųĮŌ╝░PHA║Ž│╔.R1ņoų├Č╬║═R2ģÆč§Č╬ßī┴ū┴┐(Prel)ĪóVFAöz╚Ī┴┐(VFAup)ĪóPHA║Ž│╔┴┐(PHAsyn)╝░╠ŪįŁĮĄĮŌ┴┐(Glydeg)ų«ķgĄ─▐D╗»▒╚└²║═PHAĖ„ĮMĘų░┘Ęų▒╚┼c╬─½Ił¾Ą└Ą─é„ĮyģÆč§Č╬Ą─öĄō■▒╚▌^ĮY╣¹ęŖ▒Ē 1.
ĪĪĪĪR1ņoų├Č╬PHA║Ž│╔┴┐Īó╠ŪįŁĘųĮŌ┴┐╝░ßī┴ū┴┐į┌é„ĮyģÆč§Č╬Ą─ūā╗»ĘČć·ā╚Ż¼▒Ē├„PAOsį┌ņoų├Č╬ā╚Ą─┤·ųxĘĮ╩Į┼cé„ĮyģÆč§Č╬ā╚Ą─┤·ųxŽÓ╦Ų.ė╔▒Ē 1┐╔ų¬Ż¼R1║═R2Ą─Glydeg/PHAsynĘųäe×ķ0.65Īó0.60.Arun Ą╚(1988)║═CarvalhoĄ╚(2007)šJ×ķįō▒╚└²į┌0.28~0.36ų«ķg╗“Ė³Ė▀ĢrŅA╩ŠEBPR▀\ąą│╔╣”.«öPrel/VFAup▌^ąĪ(0.15~0.45)Ż¼Č°Glydeg/VFAup▌^Ė▀(0.67~0.78)ĢrŻ¼PHVš╝PHA▒╚ųž(PHV/PHA)├„’@Ų½Ė▀(20.0%~28.6%);ŽÓĘ┤Ż¼«öPrel/VFAup▌^Ė▀(0.48~0.96)Ż¼Č°Glydeg/VFAup▌^Ą═(0.46~0.53)ĢrŻ¼PHVš╝PHA▒╚ųž(PHV/PHA)├„’@Ų½Ą═(6%~12%).ęę╦ßŌcū„×ķ╠╝į┤▒╗PAOs╬³╩š║¾Ż¼ų▒Įė▐D╗»×ķęꧯ-CoAŻ¼║¾š▀īóŠ█║Ž│╔PHB.Ą½PAOs─▄═©▀^▀ĆįŁąį╚²¶╚╦ß裣h(TCA)+ń·ńĻ╦ß-▒¹╦ß═ŠÅĮ╔·│╔▒¹§Ż-CoAŻ¼▀MČ°┼cęꧯ-CoA║Ž│╔PHV(Kortstee et al., 2000).▒ŠčąŠ┐ųąR1║═R2 Ą─Prel/VFAupŲ½Ą═Ż¼Č°Glydeg/VFAup║═PHVš╝▒╚Ų½Ė▀.─│ą®Š█┴ūŠ·┐╔─▄Ė³ČÓĄžę└┘ć╠ŪįŁĮĄĮŌŻ¼Č°▀ĆįŁąįTCA┐╔ŲĮ║Ōį┤ė┌╠ŪĮ═ĮŌ«a╔·Ą─▀ĆįŁ┴”(Oehmen et al., 2010).ō■┤╦═ŲöÓŻ¼Š█┴ūŠ·Ė³ČÓĄžę└┘ć╠ŪįŁĮĄĮŌ╠ß╣®─▄┴┐Ż¼«a╔·▌^ČÓ▀ĆįŁ┴”Ż¼ę“Č°×ķ┴╦ŲĮ║Ō▀ĆįŁ┴”Ż¼PAOs═©▀^įō═ŠÅĮ║Ž│╔▌^ČÓĄ─▒¹§Ż-CoAŻ¼▀MČ°║Ž│╔▌^ČÓĄ─PHV.ęį╔ŽĘų╬÷▒Ē├„Ż¼ņoų├Č╬─▄ŲģÆč§Č╬ū„ė├Č°╩Īģs┴╦öć░ĶŻ¼Š█┴ūŠ·─▄ā×ä▌╔·ķL▓ó┐ņ╦┘ßī┴ūŻ¼×ķ║├č§čĖ╦┘öz┴ūĄņČ©╗∙ĄA.«ö╚╗Ż¼▒ŠčąŠ┐╦∙▓╔ė├Ą─Ę┤æ¬Ų„▌^ąĪŻ¼į┌ę╗Č©│╠Č╚╔Ž└¹ė┌╬’┘|é„▀f.Č°ī”ė┌▌^┤¾Ę┤æ¬Ų„╩Ūʱėą═¼śėą¦╣¹Ż¼ęį╝░ŲõŽÓæ¬Ą─Ė─┴╝┤ļ╩®ätąĶ▀Mę╗▓Į蹊┐.
ĪĪĪĪ▒Ē1 é„ĮyģÆč§Č╬┼cņoų├Č╬Ą─╗»īWėŗ┴┐öĄ╝░PHAĮMĘų░┘Ęų▒╚Ą─▒╚▌^(ęįęę╦ßŌc×ķ╠╝į┤)
ĪĪĪĪ3.2 R1║═R2│²┴ūąį─▄▒╚▌^
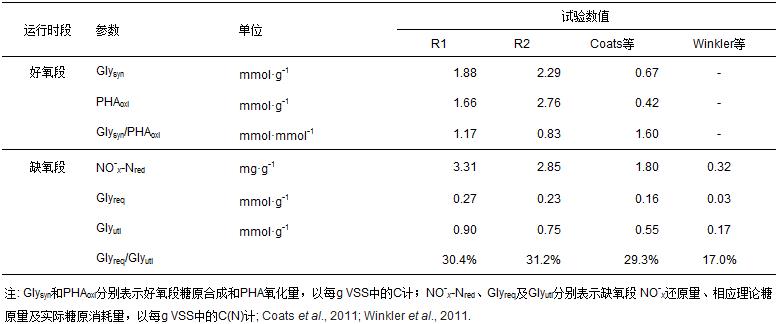
ĪĪĪĪPAOsį┌ā╔ŽĄĮy║├č§Č╬Š∙─▄čĖ╦┘╬³╩šÅU╦«ųąĄ─┴ū╦ß¹}.R1ĪóR2ųąSOPĘųäeÅ─║├č§Č╬│§Ą─46.67Īó82.34 mg Īż L-1Ž┬ĮĄĄĮ║├č§Č╬─®Ą─2.32Īó0.93 mg Īż L-1Ż¼Ųõ┴ū╦ß¹}Ą─ŲĮŠ∙╬³╩š╦┘┬╩Ęųäe×ķ5.90Īó9.53 mg Īż g-1 Īż h-1Ż¼R2Ė▀ė┌R1.R2ģÆč§Č╬Š█┴ūŠ·ßī┴ūĖ³│õĘųŻ¼ę“Č°öz┴ū╦┘Č╚Ė³┐ņŻ¼║├č§Č╬─®SOP╝┤į┌1 mg Īż L-1ęįŽ┬;Č°R1ņoų├Č╬ßī┴ū▌^R2╔┘Ż¼öz┴ū╦┘Č╚ęÓŽÓī”▌^┬².Å─║├č§┤·ųx┐┤Ż¼R1┼cR2ŽÓ═¼Ż¼ļSų°┴ūĄ─öz╚ĪR1║├č§Č╬░l╔·PHAĮĄĮŌ┼c╠ŪįŁča│õŻ¼Ą½R2Ą─Glysyn/PHAoxi▌^R1ąĪ(▒Ē 2)Ż¼ŲõįŁę“┐╔─▄╩Ūį┌R2ģÆč§Č╬ßī┴ūĖ³ČÓĄ─ŪķørŽ┬Ż¼R2ųąŠ█┴ūŠ·č§╗»PHA╦∙½@─▄┴┐▌^ČÓĄžė├ė┌SOPöz╚Ī╝░Š█┴ū║Ž│╔Ż¼Č°ė├ė┌╠ŪįŁča│õätŽÓī”▌^╔┘Ż¼╣╩Glysyn/PHAoxi▌^ąĪ.ļm╚╗R1║├č§Č╬─®SOP▌^Ė▀Ż¼×ķ2.32 mg Īż L-1Ż¼Ą½ļSų°Ę┤Ž§╗»▀MąąŻ¼SOPųØu╬³╩šŻ¼Å─Č°╩╣SOPį┌╚▒č§ĮY╩°ĢrĮĄų┴0.91 mg Īż L-1.ę“┤╦Ż¼═ŲöÓR1ųą┤µį┌Ę┤Ž§╗»Š█┴ūŠ·(DPAOs)Ż¼─▄ęįNO×ķļŖūė╩▄¾wöz╚ĪSOP(Xu et al.Ż¼ 2011)Ż¼Ę┤Ž§╗»öz┴ū╦┘┬╩×ķ0.18 mg Īż g-1 Īż h-1.├ōĄ¬Ą─═¼Ģr╩╣SOPØŌČ╚▀Mę╗▓ĮĮĄĄ═Ż¼▀_ĄĮ┼cR2ŽÓ«öĄ─│²┴ūą¦╣¹.R1║═R2ķLŲ┌▀\ąą▀^│╠ųąĄ─│²┴ūą¦╣¹ęŖłD 2.ė╔łD 2┐╔ų¬Ż¼R1║═R2ķLŲ┌▀\ąą▀^│╠ųąŻ¼│²┴ūą¦╣¹ĘĆČ©┴╝║├Ż¼ŲĮŠ∙│÷╦«ØŌČ╚Ęųäe×ķ0.91║═0.93 mg Īż L-1Ż¼ŲĮŠ∙╚ź│²┬╩Ęųäe×ķ92.4%║═92.1%.┼cįOģÆč§Č╬Ą─R2ŽÓ▒╚Ż¼įOņoų├Č╬Ą─R1Š▀ėąŽÓ«öĄ─│²┴ūą¦╣¹.▀@▒Ē├„┼cģÆč§/║├č§/╚▒č§SBRŽÓ▒╚Ż¼ņoų├/║├č§/╚▒č§SBR─▄į┌ķLŲ┌▀\ąą▀^│╠ųą╚ĪĄ├┼cų«ŽÓ«öĄ─┴╝║├│²┴ūą¦╣¹.

ĪĪĪĪłD 2 R1║═R2ķLŲ┌▀\ąąĄ─│²┴ūą¦╣¹
ĪĪĪĪ▒Ē2 ║├č§Č╬║═║¾ų├╚▒č§Č╬╬ó╔·╬’┤·ųx
ĪĪĪĪ3.3 R1║═R2├ōĄ¬ąį─▄▒╚▌^
ĪĪĪĪłD 3×ķR1║═R2Ąõą═ų▄Ų┌ā╚NH+4-NĪóNO-2-N║═NO-3-NĄ─ūā╗»Ūķør.ė╔łD 3┐╔ų¬Ż¼║├č§Č╬ā╚ā╔ŽĄĮyųąĄ─░▒Ą¬Äū║§▒╗═Ļ╚½č§╗»Ż¼ų┴║├č§Č╬─®R1║═R2ųą░▒Ą¬ØŌČ╚Ęųäe×ķ0.27Īó0.19 mg Īż L-1Ż¼╚ź│²┬╩Ęųäe×ķ99.3%║═99.5%.░▒Ą¬▒╗č§╗»│╔Ž§Ą¬╗“ü厧Ą¬Ż¼Ą½ė╔łD 3┐╔ų¬Ż¼║├č§Č╬░▒Ą¬£p╔┘┴┐┼cNO-x╔·│╔┴┐▓ŅŠÓ▌^┤¾Ż¼ę“┤╦Ż¼┐╔┐╝æ]║├č§Č╬╩Ūʱ░l╔·═¼▓ĮŽ§╗»-Ę┤Ž§╗».║├č§Č╬░▒Ą¬ų„ę¬ėą3éĆ╚źŽ“Ż║č§╗»│╔NO-xŻ¼═¼▓ĮŽ§╗»-Ę┤Ž§╗»×ķN2╝░╝Ü░¹═¼╗»×ķ╔·╬’Ą¬.╔·╬’Ą¬┐╔ęį╝Ü░¹Ęųūė╩Į C5H7NO2ėŗ╦Ń(Nš╝▒╚12%)(Bruce et al., 2001).ę“Ę┤æ¬Ų„ųą┐é╔·╬’┴┐╠Äė┌ŲĮ║ŌĀŅæBŻ¼ät├┐éĆų▄Ų┌┼┼Ę┼Ą─╬█─Ó┴┐欎ӫöė┌įōų▄Ų┌ą┬į÷Ą─╔·╬’┴┐.├┐ų▄Ų┌╗ņ║Žę║┼┼Ę┼┴┐×ķ30 mLŻ¼į┘ė╔MLVSS║═╔·╬’Ą¬š╝▒╚┐╔ėŗ╦Ń├┐ų▄Ų┌═¼╗»╔·╬’Ą¬┴┐.═¼▓ĮĘ┤Ž§╗»├ōĄ¬┴┐(SDN)ė╔▒╗č§╗»Ą─░▒Ą¬┐█│²╔·│╔Ą─NO-x║══¼╗»Ą─╔·╬’Ą¬Č°Ą├.Įøėŗ╦ŃŻ¼R1║═R2═¼▓ĮĘ┤Ž§╗»├ōĄ¬┴┐Ęųäe×ķ4.84Īó2.62 mg Īż L-1Ż¼Ęųäeš╝▀M╦«┐饬Ą─18.0%Īó9.8%(▒Ē 3).║├č§Č╬░l╔·═¼▓ĮĘ┤Ž§╗»Ą─įŁę“┐╔─▄╩Ū╗Ņąį╬█─Óą§─²¾wÅ─▒Ē├µĄĮā╚║╦Ż¼č§Ą─Ęų▓╝▓ó▓╗Š∙ä“.═Ōīė×ķ║├č§ģ^Ż¼▀MąąŽ§╗»Ę┤æ¬;ā╚īė×ķ╚▒č§ģ^Ż¼▀MąąĘ┤Ž§╗»Ę┤æ¬.SatohĄ╚(2003)░l¼FŽ§╗»║═Ę┤Ž§╗»─▄į┌ą§─²¾wĄ─▓╗═¼ģ^ė“═¼Ģr▀Mąą.┤╦═ŌŻ¼ę▓ėą┐╔─▄┤µį┌║├č§Ę┤Ž§╗»Š·(Kim et al., 2005).ļm╚╗▀M╦«░▒Ą¬ØŌČ╚ŽÓ═¼Ż¼Ą½R1═¼▓ĮĘ┤Ž§╗»├ōĄ¬┴┐▒╚R2ČÓŻ¼┐╔─▄╩Ūė╔ė┌R1ųą░▒č§╗»Š·(AOBs)ŽÓī”ü厧╦ß¹}č§╗»Š·(NOBs)Ė³ČÓŻ¼Å─Č°ü厧╦ßĘe└█┴┐Ė³ČÓ.ė╔łD 3┐╔ų¬Ż¼║├č§Č╬ā╚R1ųąü厧╦ß¹}Ęe└█┴┐┐╔▀_6.31 mg Īż L-1Ż¼Č°R2ųąü厧╦ß¹}Ęe└█┴┐ūŅČÓāH×ķ2.40 mg Īż L-1Ż¼ę“Č°Ė³ČÓĄ─ü厧╦ß¹}┐╔─▄ų▒Įė═©▀^Č╠│╠Ę┤Ž§╗»▀ĆįŁ×ķĄ¬ÜŌŻ¼Č°▓╗▒ž▀Mę╗▓Įč§╗»×ķŽ§╦ß¹}į┘ų▓Į▀ĆįŁ×ķĄ¬ÜŌ;Č°R2Ęe└█Ą─ü厧╦ß▌^╔┘Ż¼┐╔─▄ę“Ųõ▀Mę╗▓Įč§╗»×ķŽ§Ą¬Ż¼╣╩Ę┤Ž§╗»ĢrąĶŽ╚▀ĆįŁ×ķü厧Ą¬į┘▀ĆįŁ×ķĄ¬ÜŌ.ę“┤╦Ż¼R1═¼▓ĮŽ§╗»-Ę┤Ž§╗»Ė³┐ņ.
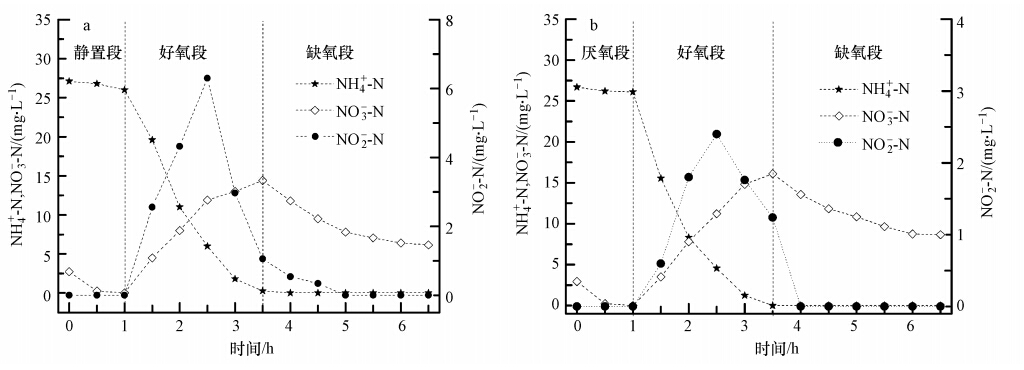
ĪĪĪĪłD 3 R1(a)║═R2(b)Ąõą═ų▄Ų┌ā╚NH+4-NĪóNO-2-N║═NO-3-NĄ─ūā╗»
ĪĪĪĪR2═Ō╠╝į┤į┌ģÆč§Č╬ęč═Ļ╚½Ž¹║─Ż¼Č°R1═Ō╠╝į┤į┌ņoų├Č╬ĮY╩°Ģrļmėą╩ŻėÓŻ¼Ą½ŲžÜŌ║¾ät┴ó╝┤╬³╩šŻ¼╝┤R1║═R2╚▒č§Č╬ęč¤o═Ō╠╝į┤.ė╔łD 1║═łD 3┐╔ų¬Ż¼R1║═R2╚▒č§Č╬ā╚NO-xųØuŽ┬ĮĄŻ¼Ūę┤╦ĢrPHA▒Ż│ųŲĮĘĆ▓ó╬┤Ž┬ĮĄŻ¼Č°╠ŪįŁätųØuŽ┬ĮĄŻ¼ę“┤╦Ż¼┐╔═ŲöÓR1║═R2į┌╚▒č§Č╬░l╔·┴╦ęįNO-x×ķļŖūė╩▄¾wŻ¼ęį░¹ā╚╠ŪįŁ×ķļŖūė╣®¾wĄ─║¾ų├Ę┤Ž§╗»Ę┤æ¬Ż¼▀@┼cCoats Ą╚(2011)║═WinklerĄ╚(2011)Ą─ł¾Ą└ę╗ų┬.╬ó╔·╬’ā╚į┤ąį╦źĮŌ“īäėĘ┤Ž§╗»╦┘┬╩×ķ0.2~0.6 mg Īż g-1 Īż h-1(Kujawa et al., 1999)Ż¼▒Š╬─ųąR1║═R2║¾ų├Ę┤Ž§╗»╦┘┬╩Ęųäe×ķ0.98║═0.84 mg Īż g-1 Īż h-1.CoatsĄ╚(2011)║═WinklerĄ╚(2011)£yĄ├║¾ų├Ę┤Ž§╗»╦┘┬╩Ęųäe×ķ0.67~0.88 mg Īż g-1 Īż h-1║═0.31~0.95 mg Īż g-1 Īż h-1(20 ĪµąŻš²Ż¼ąŻš²╣½╩ĮęŖ╬─½I(Vocks et al., 2005)).┐╔ęŖŻ¼R1║═R2║¾ų├Ę┤Ž§╗»╦┘┬╩Ė▀ė┌╬ó╔·╬’ā╚į┤ąį╦źĮŌĘ┤Ž§╗»╦┘┬╩Ż¼Č°┼c╬─½Ił¾Ą└Ą─║¾ų├Ę┤Ž§╗»╦┘┬╩ŽÓ«ö.├┐▀ĆįŁę╗Č©┴┐Ą─Ž§Ą¬ąĶŽÓæ¬ę╗Č©┴┐Ą─╠ŪįŁŻ¼2.86 mg(COD)/mg(NO-3-N)▒Ē╩Š├┐▀ĆįŁ1 mg NO-3└Ēšō╔ŽąĶ2.86 mg COD(Winkler et al.Ż¼ 2011).ō■┤╦ėŗ╦ŃŻ¼├┐▀ĆįŁ1 mg NO-2-NätąĶ1.14 mg COD.×ķĖ³║├Ąž╠Įėæ╠ŪįŁ“īäė║¾ų├Ę┤Ž§╗»├ōĄ¬Ż¼Įyėŗ┴╦Ž§Ą¬┼c╠ŪįŁų«ķgĄ─Č©┴┐ĻPŽĄöĄō■Ż¼Š▀¾węŖ▒Ē 2.Å─▒Ē 2┐╔ų¬Ż¼R1║═R2╚▒č§▀ĆįŁŽ§Ą¬└Ēšō╦∙ąĶ╠ŪįŁĘųäeš╝╚▒č§īŹļHŽ¹║─╠ŪįŁĄ─30.4%║═31.2%Ż¼▀@┼cCoats Ą╚(2011)Ą─蹊┐ĮY╣¹ŽÓ«öŻ¼Č°▌^WinklerĄ╚(2011)Ą─蹊┐ĮY╣¹Ė▀.╚▒č§Č╬▓╗ĄĮ1/2Ą─╠ŪįŁė├ė┌Ž§Ą¬▀ĆįŁŻ¼╩ŻėÓ╠ŪįŁ┐╔─▄═©▀^╠ŪĮ═ĮŌ×ķ╬ó╔·╬’╣®─▄ęįŠS│ų╔·ķL.R1╚▒觎¹║─╠ŪįŁ▌^R2ČÓŻ¼ŲõŽ§Ą¬▀ĆįŁ┴┐ęÓ▌^ČÓ(▒Ē 2).R1Ę┤Ž§╗»╦┘┬╩Ė▀ė┌R2Ż¼┐╔─▄Š═╩Ūė╔ė┌R1╚▒č§Č╬ĘųĮŌĄ─╠ŪįŁ┴┐(0.90 mmol Īż g-1)▌^R2(0.75 mmol Īż g-1)ĘųĮŌĄ─ČÓ(▒Ē 2).ų┴ė┌×ķ║╬R1╚▒č§╠ŪįŁĘųĮŌ┴┐▌^R2ČÓŻ¼ŲõįŁę“┐╔─▄╩ŪR2 ║├č§Č╬č§╗»PHA╦∙½@─▄┴┐Ė³ČÓĄžė├ė┌SOPöz╚Ī╝░ā”┤µŻ¼ė├ė┌╠ŪįŁča│õätŽÓī”▌^╔┘Ż¼Å─Č°ī¦ų┬╚▒č§Č╬╠ŪįŁĘųĮŌ▌^╔┘.ė╔▒Ē 2▀Ć┐╔ų¬Ż¼R1ųąGlysyn/PHAoxi▌^R2ąĪ.R1║═R2╚▒č§Č╬Ę┤Ž§╗»╦┘┬╩(DNR)╚ń▒Ē 4╦∙╩Š.Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.bnynw.comĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
ĪĪĪĪ▒Ē3 R1║═R2║├č§Č╬═¼▓ĮŽ§╗»-Ę┤Ž§╗»

ĪĪĪĪ▒Ē4 R1║═R2╚▒č§Č╬Ę┤Ž§╗»╦┘┬╩
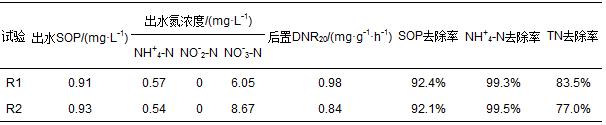
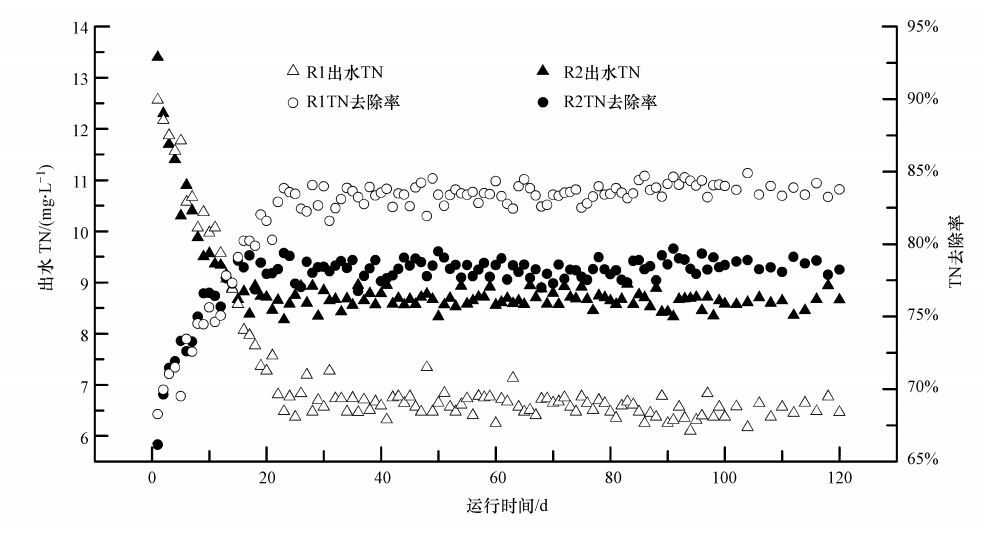
ĪĪĪĪŠC║Žī”▒╚R1║═R2║├č§Č╬║═╚▒č§Č╬├ōĄ¬ąį─▄░l¼FŻ¼įOų├ņoų├Č╬Ą─R1▒╚įOų├ģÆč§Č╬Ą─R2ėąĖ³Ė▀Ą─═¼▓ĮŽ§╗»-Ę┤Ž§╗»├ōĄ¬┴┐║═Ę┤Ž§╗»╦┘┬╩Ż¼╣╩į┌░▒Ą¬č§╗»┴┐Äū║§ŽÓ═¼ŪķørŽ┬Ż¼R1Ž§Ą¬│÷╦«ØŌČ╚▌^R2Ą═(▒Ē 4).ę“Č°Š▀ėąĖ³║├Ą─├ōĄ¬ą¦╣¹.R1║═R2ķLŲ┌▀\ąą▀^│╠ųą├ōĄ¬ą¦╣¹ęŖłD 4.ė╔łD 4┐╔ų¬Ż¼ķLŲ┌▀\ąą▀^│╠ųąR1║═R2├ōĄ¬ą¦╣¹ĘĆČ©Ż¼ŲĮŠ∙│÷╦«TNĘųäe×ķ6.62║═9.21 mg Īż L-1Ż¼ŲĮŠ∙├ōĄ¬┬╩Ęųäe×ķ83.5%║═77.0%.┐╔ęŖŻ¼R1├ōĄ¬ą¦╣¹║├ė┌R2.
ĪĪĪĪłD 4 R1║═R2ķLŲ┌▀\ąąĄ─├ōĄ¬ą¦╣¹
ĪĪĪĪ4 ĮYšō
ĪĪĪĪį┌ā╔Ę┤æ¬Ų„▀M╦«ęę╦ßŌcĪó░▒Ą¬(NH+4-N)╝░┴ū╦ß¹}(PO3-4-P)ØŌČ╚Š∙Ęųäe×ķ350 mg Īż L-1(ęįCODėŗ)Īó40 mg Īż L-1╝░12 mg Īż L-1Ż¼╦«┴”═Ż┴¶Ģrķg(HRT)×ķ12 hĢrŻ¼ī”▒╚蹊┐┴╦ņoų├/║├č§/╚▒č§SBR(R1)┼cģÆč§/║├č§/╚▒č§SBR(R2)Ą─╔·╬’├ōĄ¬│²┴ūąį─▄. ĮY╣¹░l¼FŻ¼ņoų├Č╬┐╔ŲģÆč§Č╬ū„ė├Ż¼▀@×ķ╔·╬’ÅŖ╗»│²┴ūĄņČ©┴╦╗∙ĄA;ņoų├/║├č§/╚▒č§SBRĄ─ SOP║═TN╚ź│²┬╩Ęųäe×ķ92.4%║═83.5%Ż¼ģÆč§/║├č§/╚▒č§SBRĄ─SOP║═TN╚ź│²┬╩Ęųäe×ķ92.1%║═77.0%;ęįņoų├Č╬┤·╠µģÆč§Č╬Ą─ņoų├/║├č§/╚▒č§SBR┐╔▀_ŽÓ«öĄ─│²┴ūą¦╣¹Ż¼Č°ė╔ė┌R1═¼▓ĮŽ§╗»-Ę┤Ž§╗»├ōĄ¬┴┐╝░║¾ų├Ę┤Ž§╗»├ōĄ¬┴┐Š∙▌^R2ČÓŻ¼╣╩Ųõ├ōĄ¬ą¦╣¹Ė³║├.įōįOų├į┌▒▄├Ō╬█─Ó╗ž┴„Ą─╗∙ĄA╔Ž╩ĪģsģÆč§öć░ĶČ°║å╗»╣ż╦ćŻ¼ĮĄĄ═┴╦╠Ä└Ē│╔▒Š.

