Bų│╬█╦«╠Ä└Ē╣ż╦ć
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2017-9-14 9:31:27
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ└¹ė├╬óįÕ▀Mąą╬█╦«╠Ä└ĒĄ─Üv╩ĘūĘ╦▌ęčŠ├. įńį┌20╩└╝o50─Ļ┤·Ż¼OswaldĄ╚Š═╠ß│÷└¹ė├╬óįÕ╠Ä└Ē╬█╦«Ą─įOŽļ. ┤╦║¾Ż¼ęįįÕ-Š·╣▓╔·¾wŽĄ║═Ė▀ą¦įÕŅÉ╠┴×ķ┤·▒ĒĄ─æęĖĪ╔·ķLįÕŅÉ╠┴ŽĄĮyį┌Ęų╔ó╩Į╬█╦«╠Ä└ĒųąĄ├ĄĮ┴╦ÅVĘ║Ą─╣ż│╠æ¬ė├. Ą½▀@ŅÉŽĄĮyę“š╝Ąž├µĘe┤¾Īó╠Ä└Ēą¦╣¹▓╗ĘĆČ©Ą╚ŠųŽ▐ąįŻ¼ę╗ų▒╬┤─▄│╔×ķ╬█╦«╠Ä└ĒĄ─ų„┴„╣ż╦ć. Į³─ĻüĒŻ¼į┌╩ąš■╬█╦«╠Ä└ĒÅS╔ŅČ╚ā¶╗»ąĶę¬ęį╝░┐╩═¹Å─╬█╦«ųą½@Ą├╔·╬’▓±ė═Ą─“īäėŽ┬Ż¼╬óįÕ╬█╦«╠Ä└Ēį┌╩└ĮńĘČć·ā╚ųž½@ą┬╔·.
ĪĪĪĪ╬óįÕ╔·ķL▀^│╠ąĶę¬┤¾┴┐╬³╩šĄ¬(N)Īó┴ū(P)Ą╚ĀIBį¬╦žŻ¼┐╔ų▒ĮėĮĄĄ═Č■/╚²╝ē│÷╦«ųąNĪóPĄ╚╬█╚Š╬’Ą─║¼┴┐. ═©▀^╣╠Č©Č■č§╗»╠╝(CO2)Īó«a╔·č§ÜŌ(O2)Īó╠ßĖ▀pHĄ╚ķgĮėū„ė├Ż¼╬óįÕ▀Ć─▄äōįņ│÷ėąą¦╚ź│²╦«ųąÜł┴¶ėąÖC╬’║═▓ĪįŁąį╬ó╔·╬’Ą─ŁhŠ│Śl╝■. ┤╦═ŌŻ¼╬óįÕę▓Š▀ėą╬³ĖĮųžĮī┘Ą╚ėą║”╬’┘|Ą──▄┴”. ę“┤╦Ż¼╬óįÕŠ▀ėą│╔×ķ╬█╦«╔ŅČ╚ā¶╗»╝╝ągĄ─┴╝║├Øō┴”. į┌╬█╦«Č■/╚²╝ē╠Ä└ĒųąŻ¼╚ź│²ĀIBį¬╦žĄ─│ŻęŖįÕĘN░³└©: ó┘ŠGįÕķTĄ─ąĪŪ“įÕ(Chlorella)ĪóŲŽ╠čįÕ(Botryococcus)Īó¢┼įÕ(Scenedesmus)║═╬óŠGŪ“įÕ(Nannochloris)Ą╚Ż¼Ųõųąė╚ęįąĪŪ“įÕ(Chlorella)║═¢┼įÕ(Scenedesmus)Ą─蹊┐ł¾Ą└×ķČÓ;ó┌╦{įÕķTĄ─╣Øą²įÕī┘(Arthrospira sp.)ĪóŅØįÕī┘(Oscillatoria sp.)║═Ž»įÕī┘(Phormidium);ó█╣ĶįÕķTĄ─╚²ĮŪ║ųųĖįÕ(P. tricornutum)Ą╚.ęį╔ŽĘNī┘Ą─NĪóP╚ź│²ą¦╣¹┐╔ģóęŖCaiĄ╚Ą─ŠC╩÷╬─š┬. į┌įÕĘN▀xō±Ą─╗∙ĄA╔ŽŻ¼╬óįÕ┼ÓBŽĄĮy(Ę┤æ¬Ų„)Ą─śŗĮ©╩ŪīŹ¼F╬óįÕ╬█╦«╠Ä└Ē╣ż│╠╗»æ¬ė├Ą─ĻPµI. ░┤╬óįÕĄ─╔·ķLĘĮ╩Į▓╗═¼Ż¼╬óįÕ┼ÓBŽĄĮy┐╔Ęų×ķæęĖĪ┼ÓB║═ĖĮų°┼ÓBā╔┤¾ŅÉ. æęĖĪ┼ÓBŽĄĮy┐╔▀Mę╗▓ĮĘų×ķķ_Ę┼╩Į║═ĘŌķ]╩Įā╔ŅÉŻ║ó┘ķ_Ę┼╩ĮŽĄĮyų„ę¬ųĖĖ„ŅÉ╠┴ŽĄĮyŻ¼Ąõą═Ą─╚ńĖ▀ą¦įÕŅÉ╠┴║═┼▄Ą└╩ĮįÕŅÉ╠┴Ą╚;ó┌ĘŌķ]╩ĮŽĄĮyų„ę¬ųĖĖ„ŅÉ╣Ō╔·╬’Ę┤æ¬Ų„Ż¼Ęų×ķ╣▄╩Į(┤╣ų▒Īó╦«ŲĮĪó┬▌ą²)ĪółAų∙╩ĮĪó▒Ī░Õ╩Į║═┤³╩ĮĄ╚. ĖĮų°╩ĮŽĄĮy░³└©╣Ō╔·╬’─ż(ŲĮ░Õ)Ę┤æ¬Ų„║═įÕ╝Ü░¹╣╠Č©╗». ┐╝æ]ĄĮ╬█╦«╠Ä└ĒĄ─īŹļHŪķør(╦«┴┐┤¾Ż¼Į©įņĪó▀\ąą│╔▒ŠĄ╚)Ż¼ķ_Ę┼┼ÓBŽĄĮy╚įīó╩Ū╬óįÕ╬█╦«╠Ä└ĒĄ─ų„┴„Ę┤æ¬Ų„śŗą═.
ĪĪĪĪ╚ń╔Ž╦∙╩÷Ż¼įÕ╝Ü░¹ė├ė┌╔·«a╔·╬’▓±ė═╩Ū╬óįÕ╬█╦«╠Ä└Ēųž½@ą┬╔·Ą─ų„ꬓīäė┴”ų«ę╗. ═©▀^╬óįÕ╔·«a╔·╬’▓±ė═Š▀ėąŲõ╦¹╚╬║╬«aė═ū„╬’¤oĘ©▒╚öMĄ─ā×ä▌Ż║ó┘įÕ╝Ü░¹Ą─╣Ō║Žą¦┬╩Ė▀Ż¼╔·ķL╦┘Č╚┐ņĪóų▄Ų┌Č╠Ż¼Ųõ«aė═┴┐×ķ47000~190000 L Īż hm-2 Īż a-1Ż¼╩Ū▐rū„╬’Ą─7~30▒Č;ó┌╔·╬’┘|╚╝ė═¤ßųĄĖ▀Ż¼ŲĮŠ∙▀_33 MJ Īż kg-1Ż¼╩Ū─Š▓─╗“▐rū„╬’ĮšČÆĄ─1.6▒Č;ó█▓╗ąĶš╝ė├▐rśIė├Ąž;ó▄╔·╬’┘|(įÕ╝Ü░¹)╔·«a║═╝ė╣ż│╔▒ŠĄ═Ż¼ė╚Ųõ╩Ūęį╬█╦«×ķĄū╬’▀MąąįÕ╝Ü░¹┼ÓBĢr. ėąĶbė┌┤╦Ż¼├└ć°ĪóÜWų▐Īó░─┤¾└¹üåĪó╚š▒ŠĪó┼_×│Ą╚░l▀_ć°╝ę║═Ąžģ^Č╝ęčīó╬óįÕ┼ÓBū„×ķīŹ¼F╬█╦«╔·æB╠Ä└Ē║═┐╔į┘╔·─▄į┤╔·«aĄ─æ┬į░lš╣─┐ś╦. │ŻęŖĄ─«aė═įÕĘN╝░Ųõė═ų¼║¼┴┐╬─½Ięč╩÷╝░. ╣żśI╔Žęį«aė═×ķ─┐Ą─Ą─╬óįÕ┼ÓBę╗░Ń▓╔ė├ĘŌķ]╩Į╣Ō╔·╬’Ę┤æ¬Ų„Ż¼▓óŪę═∙═∙▓╔ė├╝ā┼ÓB╗“å╬ųĻ┼ÓBĄ─ĘĮ╩Į. «öĮY║Ž╬█╦«╠Ä└Ē─┐ś╦ĢrŻ¼ę“Š▐┤¾Ą─╦«┴┐╝░╬█╦«ųąÅ═ļsĄ─│╔Ęų(ė╚Ųõ╩ŪŲõųą░³║¼Ą─╗ņ║ŽĘNī┘)Ż¼ęį╔Ž┼ÓBĘĮ╩Įīó║▄ļyŠS│ų.
ĪĪĪĪĮ³─ĻüĒŻ¼ć°ā╚═ŌīWš▀į┌ķ_░l╬óįÕ╬█╦«╔ŅČ╚ā¶╗»║═┐╔į┘╔·─▄į┤╔·«aØō┴”ĘĮ├µ▀Mąą┴╦┤¾┴┐蹊┐;į┌╬█╦«ā¶╗»ÖC└ĒĪóįÕĘN║Y▀xĪóĘ┤æ¬Ų„įOėŗĪó╣ż╦ćŚl╝■┐žųŲ╝░įÕ╝Ü░¹╝ė╣ż└¹ė├Ą╚ĘĮ├µČ╝╚ĪĄ├┴╦ĘeśOĄ─▀Mš╣. ╚╗Č°Ż¼¤ošōÅ─╬█╦«ā¶╗»▒Š╔ĒŻ¼▀Ć╩Ū─▄į┤╔·«aüĒšfŻ¼įÕ╝Ü░¹Ą─ĘųļxĪó▓╔╩šČ╝ę╗ų▒╩Ūę╗éĆæęČ°╬┤øQĄ─╗∙ĄAąį╝╝ągļyŅ}. ╬óįÕ╝Ü░¹ę╗░ŃąĪė┌30 ”╠mŻ¼Ä¦žōļŖ║╔Ż¼├▄Č╚ĮėĮ³ė┌╦«Ż¼▀@ą®╠žąį╩╣Ą├įÕ╝Ü░¹į┌╦«ųą═∙═∙╠Äė┌ĘĆČ©Ą─æęĖĪĀŅæBŻ¼║▄ļyŽ±╗Ņąį╬█─Ó─Ūśė═©▀^ųž┴”│┴ĄĒČ°īŹ¼Fūį╚╗Ęųļx. ĮY╣¹Ż¼įÕ╝Ü░¹Ģ■ļS╠Ä└Ē╦«┤¾┴┐┴„╩¦Ż¼▓╗āHČ■┤╬╬█╚Š╠Ä└Ē╦«Ż¼Č°Ūęī¦ų┬Ę┤æ¬Ų„ā╚╔·╬’┴┐ļyęį┤¾┴┐ŠS│ų(ę╗░ŃāH×ķ0.2~0.6 g Īż L-1). Ą═Ą─┼ÓB├▄Č╚ī¦ų┬╚ź│²ą¦┬╩Ą═Ž┬Ż¼╩╣Ą├╠Ä└Ēą¦╣¹ĘĆČ©ąį▌^▓Ņ. ī”┤╦Ż¼═∙═∙ąĶĮĄĄ═╠Ä└Ēžō║╔Ż¼═¼Ģr▓╔ė├▌^ķLĄ─╦«┴”═Ż┴¶Ģrķg(HRT)Ż¼▀MČ°ī¦ų┬š╝Ąž├µĘe╝ė┤¾. ─┐Ū░Ųš▒ķæ¬ė├Ą─įÕŅÉ╠┴ŽĄĮyHRTę╗░Ń×ķ2~6 dŻ¼«ö┴┐╚╦┐┌š╝Ąžę╗░Ń>10 m2. ’@╚╗Ż¼Ųõš╝Ąž├µĘeę¬▒╚Č■/╚²╝ē╬█╦«╠Ä└Ēų„¾wå╬į¬▀Ćę¬²ŗ┤¾įSČÓŻ¼▀@į┌ė├ĄžŠoÅłĄ─│Ū╩ąųą╩Ū║▄ļy▒╗Įė╩▄Ą─.
ĪĪĪĪÅ──▄į┤╔·«aĮŪČ╚┐┤Ż¼ØMūŃ╣żśI└¹ė├ę¬Ū¾Ą─įÕ╝Ü░¹įŁ┴ŽŲõūŅ╝č╔·╬’┴┐æ¬▀_ĄĮ300~400 g Īż L-1(Ė╔┘|┴┐). ę“┤╦Ż¼│ŻęÄ┼ÓBŽ┬Ą─įÕę║ąĶØŌ┐s1 000▒Čęį╔Ž║¾ĘĮ─▄į┌╣żśI╔Ž╝ėęį└¹ė├. ▀@ę╗Ė▀─▄║─Ą─ĘųļxĪóØŌ┐s▀^│╠╩Ū╬óįÕ─▄į┤╔·«aųąĄ─ų„ę¬─▄║─│╔▒Š(š╝╬óįÕ╔·╬’┘|╔·«a┐é│╔▒ŠĄ─20%~50%. ▀^Ė▀Ą─╔·«a│╔▒Š╩╣Ą├įÕŅÉ╔·«a╔·╬’▓±ė═┼c╗»╩»╚╝┴ŽŽÓ▒╚╚į╠Äė┌┴ėä▌.
ĪĪĪĪ┐╔ęŖŻ¼įÕ╝Ü░¹ĘųļxĪó▓╔╩š└¦ļy╩ŪŽ▐ųŲ╬óįÕ╝╝ąg┤¾ęÄ─Ż╣żśI╗»æ¬ė├Ą─ųžę¬Ų┐Ņi. ╬óįÕĘųļxĪó▓╔╩š│Żė├Ą─ĘĮĘ©░³└©ļxą─Ę©Īó▀^×VĘ©(░³└©─ż×V)ĪóÜŌĖĪĘ©Īóų▒Įėųž┴”│┴ĮĄĘ©║═ą§─²Ę©Ą╚. ļxą─Ę©╩Ū┐ņ╦┘Īó┐╔┐┐Ą─Ęųļx▓╔╩šĘĮĘ©. Ą½ė╔ė┌ŲõśOĖ▀Ą──▄║─║══Č┘Y▀\ąą│╔▒ŠŻ¼į┌─┐Ū░╝╝ągŚl╝■Ž┬▓ó▓╗Š▀éõ┤¾ęÄ─Ż╣ż│╠æ¬ė├Ą─Øō┴”. ▀^×VĘ©āHį┌ĘųļxĮzĀŅįÕĢr─▄║─║═│╔▒Š▌^Ą═; ī”ė┌ĘŪĮzĀŅįÕśOęūą╬│╔─ż╬█╚ŠŻ¼─▄║─║═▀\ąą│╔▒Š║▄Ė▀Ż¼▓╗─▄ØMūŃĖ▀ą¦ĪóĄ═│╔▒Š▓╔╩šĄ─ę¬Ū¾. ÜŌĖĪĘ©āH▀mė├ė┌▓╔╩šå╬╝Ü░¹įÕŅÉŻ¼į┌╬█╦«╗ņ║Ž┼ÓBĄ─Śl╝■Ž┬▓╗─▄Ųš▒ķ▀mė├; ┤╦═ŌŻ¼ė╔ė┌ꬫa╔·┤¾┴┐Ą─╬óąĪÜŌ┼▌Ż¼Ųõ═Č┘Y║═▀\ąą│╔▒Š/─▄║─ęÓ║▄Ė▀Ż¼╔§ų┴┐╔─▄Ė▀▀^ļxą─Ę©. ų▒Įėųž┴”│┴ĮĄĘ©╩Ū│╔▒ŠūŅ×ķĄ═┴«Ą─ĘųļxĪó▓╔╩šĘĮĘ©. Ą½Ųõ║─ĢrķLŻ¼Ęųļxą¦╣¹║═┐╔┐┐ąįūŅ▓Ņ.
ĪĪĪĪą§─²Ę©╩ŪĘųļx╦«ųą┤ųĘų╔ó║═─z¾w╬’┘|æ¬ė├ūŅ×ķÅVĘ║Ą─ĘĮĘ©Ż¼į┌20╩└╝o80─Ļ┤·Š═ęčĮøė├ė┌╬óįÕĄ─Ęųļx▓╔╩š(. æęĖĪįÕę║Įøą§─²║¾─▄īŹ¼FĖ▀ą¦ųž┴”│┴ĄĒĘųļx; ĘųļxĄ─įÕ╝Ü░¹─▄ų▒Įė▒╗Įž┴¶į┌Ę┤æ¬Ų„ā╚Ż¼▀_ĄĮŠS│ųĖ▀╔·╬’┴┐║═▒ŻšŽ│÷╦«╦«┘|Ą──┐Ą─. Å─å╬╝āĄ─įÕ╝Ü░¹▓╔╩šĮŪČ╚üĒšfŻ¼ą§─²Ę©╩Ū╠Ä└Ē┤¾┴┐ŽĪįÕę║ĢrūŅ×ķĮøØ·Īó┐╔ąąĄ─ĘĮĘ©. ļm╚╗įÕ╝Ü░¹Įøą§─²│┴ĄĒ║¾▀Ć▓╗─▄ų▒Įė▀_ĄĮ╣żśIæ¬ė├Ą─ę¬Ū¾Ż¼Ą½ęč─▄’@ų°ĮĄĄ═║¾└mØŌ┐s▀^│╠Ą──▄║─║═│╔▒Š. ę“┤╦Ż¼ą§─²Ę©ęč▒╗ęĢ×ķīŹ¼F╬óįÕ┤¾ęÄ─ŻĘųļx▓╔╩šĄ─ūŅ╝čĘĮĘ©. Ė∙ō■╩ŪʱąĶę¬╠Ē╝ėą§─²ä®┐╔Ęų×ķĪ░═Ō╝ėą§─²ä®Ę©Ī▒║═Ī░ūį░ląįą§─²Ę©Ī▒ā╔┤¾ŅÉ. ŲõųąŻ¼═Ō╝ėą§─²ä®Ę©Ė∙ō■╦∙╩╣ė├Ą─ą§─²ä®ĘNŅÉėų┐╔Ęų×ķ¤oÖCą§─²ä®Ę©ĪóėąÖCĖ▀Ęųūėą§─²ä®Ę©║═╔·╬’ą§─²ä®Ę©. ūį░ląįą§─²░┤šš░l╔·ÖC└Ē┐╔▀Mę╗▓ĮĘų×ķĖ▀pHšTī¦Ą─ūį░ląįą§─²║═░¹═ŌŠ█║Ž╬’(EPS)ę²ŲĄ─ūį░ląįą§─².
ĪĪĪĪ▒Š╬─īóÅ─ī”╬óįÕ▒Ē├µ╠žąį║═ą§─²ÖC└ĒĄ─║åę¬ĮķĮB│÷░lŻ¼ŽĄĮy┐éĮYĖ„ĘNą§─²ĘųļxĘĮĘ©Ą─蹊┐æ¬ė├¼FĀŅŻ¼▀MČ°ī”Ė„ĘNĘĮĘ©▀MąąŠC║Ž▒╚▌^Ż¼ęįŲ┌ūŅĮK├„┤_╬óįÕą§─²ĘųļxĄ─░lš╣ĘĮŽ“.
ĪĪĪĪ2 ╬óįÕ▒Ē├µ╠žąį║═ą§─²ÖC└Ē 2.1 ╬óįÕæęĖĪę║Š█╝»ĘĆČ©ąįĄ─└Ēšō┐“╝▄
ĪĪĪĪöUš╣DLVO(XDLVO)└Ēšō╩Ū─z¾w╗»īWųą├Ķ╩÷─z¾wĘĆČ©ąįĄ─ĮøĄõ└Ēšōų«ę╗Ż¼ęč│╔╣”æ¬ė├ė┌├Ķ╩÷╗Ņąį╬█─ÓŽĄĮy╬ó╔·╬’╝Ü░¹ķgĄ─š│ĖĮŠ█╝»(ą§─²)▀^│╠. ūŅĮ³čąŠ┐ūCīŹŻ¼įō└Ēšō═¼śė▀mæ¬ė┌├Ķ╩÷╬óįÕæęĖĪę║ųąįÕ╝Ü░¹Ą─Š█╝»▀^│╠. į┌XDLVO└ĒšōųąŻ¼─z┴ŻķgĄ─ŽÓ╗źū„ė├ų„ę¬┐╝æ]┴╦ęįŽ┬3ĘNĘŪ╣▓ārµIĄ─ŽÓ╗źū„ė├┴”Ż║ó┘ĘČĄ┬╚A┴”(Lifshitz-van der Waals interaction)Ż¼╦³╩Ū╔½╔ó┴”ĪóśOąį┴”║═šTī¦┼╝śO┴”ų«║═; ó┌ņoļŖ┴”(Electrostatic interaction)Ż¼į┤ūį─z┴Ż▒Ē├µ╦∙ĦļŖ║╔Ą─ņoļŖŽÓ╗źū„ė├; ó█Lewis╦ß-ēA╦«║Žū„ė├┴”(Lewis acid-base interaction)Ż¼į┤ūįśOąįĮMĘųķgĄ─ļŖūė▐DęŲ. ─z┴ŻķgĄ─┐é▒Ē├µ╬╗─▄(GTOT(d))×ķęį╔Žū„ė├┴”Ą─╬╗─▄ų«║═Ż║
ĪĪĪĪ╩ĮųąŻ¼GLW(d)×ķĘČĄ┬╚Aū„ė├┴”╬╗─▄Ż¼GEL(d)×ķņoļŖū„ė├┴”╬╗─▄Ż¼GAB(d)×ķLewis╦ß-ēA╦«║Žū„ė├┴”╬╗─▄.(d)▒Ē╩Šū„ė├┴”Ą─┤¾ąĪ║═ąį┘|×ķ─z┴ŻķgŠÓĄ─║»öĄ. └Ēšō╔ŽŻ¼GTOT(d)>0ät─z┴ŻķgŽÓ╗ź┼┼│ŌŻ¼╠Äė┌Š█╝»ĘĆČ©ĀŅæB; GTOT(d)<0ät─z┴ŻŽÓ╗źŠ█╝». Ąõą═Ą─┐é╬╗─▄Ū·ŠĆę╗░Ń░³║¼ā╔éĆĄ═╬╗č©─▄(─z┴ŻķgŠÓė╔▀h╝░Į³Ęųäe×ķĄ┌Č■Ą═╬╗č©─▄Em2║═Ą┌ę╗Ą═╬╗č©─▄Em1)Ż¼ā╔š▀ų«ķg┤µį┌ę╗│Ō┴”─▄ĘÕ(Eb). «ö─z┴ŻŽÓ╗ź┐┐Į³Ż¼ĄĮ▀_Ą┌Č■Ą═╬╗č©─▄³c(Em2)ĢrŻ¼─z┴Żķg╠Äė┌ę╗ĘN┐╔─µĄ─š│ĖĮĀŅæB;═ŌĮńŚl╝■╔įėąūā╗»ätš│ĖĮĄ──z┴ŻėųīóŽÓ╗źĘųļxŻ¼╩Ūę╗ĘN▓╗└╬╣╠Ą─š│ĮYĀŅæB. ų╗ėą─z┴ŻĄ─äė─▄ūŃē“┤¾Ż¼ūŃęį┐╦Ę■│Ō┴”─▄ĘÕĄĮ▀_Ą┌ę╗Ą═╬╗č©─▄(EEm1)Ģr▓┼─▄ą╬│╔└╬╣╠Ą─š│ĮYĀŅæBŻ¼╝┤░l╔·ą§─²)
ĪĪĪĪ2.2 įÕ╝Ü░¹▒Ē├µ╠žąį┼cŠ█╝»ĘĆČ©ąį
ĪĪĪĪøQČ©┐é▒Ē├µ╬╗─▄(╩Į(1))Ą─3ĘN╗∙▒Šū„ė├┴”ųąŻ¼ĘČĄ┬╚A┴”ę╗░Ń▒Ē¼F×ķę²┴”Ż¼Ųõ┤¾ąĪ╚ĪøQė┌─z┴ŻķgŠÓĪóå╬╬╗¾wĘeā╚Ą─┴ŻūėöĄ┴┐║═║═┴ŻūėĄ─śO╗»┬╩Ą╚. Č°ņoļŖ┴”║═Lewis╦ß-ēA╦«║Žū„ė├┴”Ą─ąį┘|║═┤¾ąĪät╚ĪøQįÕ╝Ü░¹Ą─▒Ē├µļŖä▌║═ėH/╩Ķ╦«ąįĄ╚▒Ē├µ╠žąį.
ĪĪĪĪ2.2.1 ▒Ē├µļŖä▌
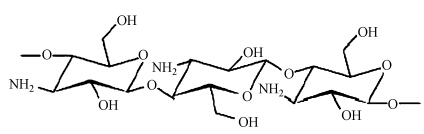
ĪĪĪĪįÕ╝Ü░¹▒Ē├µĖ▓╔wų°ę╗īėÅ═ļsĄ─EPSŻ¼Ųõų„ę¬│╔Ęų×ķ╠╝╦«╗»║Ž╬’(EPSC)║═Ą░░ū┘|(EPSP)Ż¼Ųõ╦¹│╔Ęų░³└©Ė»ų│┘|(Humus-like Substances)Īó║╦╦ß(Nucleic Acids)Īó╠Ū╚®╦ß(Uronic Acids)Ą╚. ▀@ą®│╔Ęųī¦ų┬įÕ╝Ü░¹▒Ē├µĖ╗╝»┴╦┤¾┴┐¶╚╗∙(Ī¬COOH)║═░▒╗∙(Ī¬NH2)Ą╚╣”─▄łF. ▀@ą®╣”─▄łFļS¾wŽĄpH▓╗═¼─▄Įė╩š╗“╩¦╚ź┘|ūė(H+)Ż¼ė╔┤╦ą╬│╔▒Ē├µļŖ║╔╝░ļŖä▌. └²╚ń: «ö¾wŽĄ╠Äė┌Ą═pHŚl╝■ĢrŻ¼¶╚╗∙║═░▒╗∙Č╝īóĮė╩šH+(┘|ūė╗»Ż¼protonation)Ż¼ą╬│╔š²Ą─▒Ē├µļŖ║╔;ŽÓĘ┤Ż¼«ö¾wŽĄ╠Äė┌Ė▀pHŚl╝■ĢrŻ¼¶╚╗∙īó╩¦╚źH+(╚ź┘|ūė╗»Ż¼deprotonation)Ż¼ą╬│╔žōĄ─▒Ē├µļŖ║╔;į┌╠žČ©pHŚl╝■Ž┬Ż¼┐╔ęįą╬│╔¶╚╗∙╩¦H+Č°░▒╗∙Ą├H+Ą─ŪķørŻ¼▒Ē├µā¶ļŖ║╔×ķ┴ŃŻ¼╝┤Ą╚ļŖ³c. ī”ė┌╬óįÕŻ¼ŲõĄ╚ļŖ³cę╗░Ńį┌pH=3. Č°īŹļH╬óįÕ┼ÓBŽĄĮyĄ─pHę╗░Ńį┌7ęį╔Ž. ╦∙ęįŻ¼įÕ╝Ü░¹ę╗░ŃĦžōļŖŻ¼╝┤╩Į(1)ųąĄ─ņoļŖū„ė├┴”ĒŚ▒Ē¼F×ķ│Ō┴”.
ĪĪĪĪ─z┴Ż▒Ē├µļŖä▌¤oĘ©ų▒Įė£y┴┐Ż¼ų╗─▄£y┴┐│÷─z┴ŻĄ─ZetaļŖ╬╗║¾═©▀^ėŗ╦ŃķgĮėĄ├│÷. ZetaļŖ╬╗╩Ū─z┴ŻļpļŖīėĮYśŗųą╗¼äė├µ┼c╦«╚▄ę║ų«ķgĄ─ļŖ╬╗▓ŅŻ¼╩Ū▒Ēš„Ęų╔ó¾wŽĄĘĆČ©ąįĄ─ųžę¬ųĖś╦. ZetaļŖ╬╗Į^ī”ųĄįĮĖ▀Ż¼─z┴Żų«ķgĄ─┼┼│Ō┴”įĮ┤¾Ż¼¾wŽĄįĮĘĆČ©. īŹļH┼ÓBŚl╝■Ž┬įÕŅÉĄ─ZetaļŖ╬╗ę╗░Ńį┌-35~-15 mVų«ķg. ę“┤╦Ż¼įÕ╝Ü░¹ķgĄ─ņoļŖ│Ō┴”ę╗░Ń▌^┤¾Ż¼╩ŪįÕ╝Ü░¹į┌╦«╚▄ę║ųą▒Ż│ųŠ█╝»ĘĆČ©ąįĄ─ų„ę¬įŁę“.
ĪĪĪĪ2.2.2 ėH/╩Ķ╦«ąį
ĪĪĪĪįÕ╝Ü░¹Ą─▒Ē├µėH/╩Ķ╦«ąįøQČ©┴╦╩Į(1)ųąLewis╦ß-ēA╦«║Žū„ė├┴”Ą─ąį┘|║═┤¾ąĪŻ¼Š▀¾wėą╚ńŽ┬ęÄ┬╔: ╩Ķ╦«ąįįÕ╝Ü░¹ķgĄ─Lewis╦ß-ēA╦«║Žū„ė├┴”▒Ē¼F×ķę²┴”; ėH╦«ąįįÕ╝Ü░¹ķgĄ─Lewis╦ß-ēA╦«║Žū„ė├┴”▒Ē¼F×ķ│Ō┴”; ėH╦«║═╩Ķ╦«įÕ╝Ü░¹ķgĄ─Lewis╦ß-ēA╦«║Žū„ė├┴”ąį┘|ät╚ĪøQė┌įÕ╝Ü░¹Ą─ŽÓī”ėH/╩Ķ╦«│╠Č╚Ż¼┐╔×ķę²┴”╗“│Ō┴”; ėH/╩Ķ╦«│╠Č╚įĮĖ▀Ż¼Lewis╦ß-ēA╦«║Žū„ė├┴”Ą─ųĄįĮ┤¾. ęį╔ŽęÄ┬╔┐╔═©╦ūĄž└ĒĮŌ×ķ╩Ķ╦«╝Ü░¹į┌╦«╚▄ę║ųąīó╩▄ĄĮ╦«ĘųūėĄ─Ī░┼┼│ŌĪ▒ū„ė├Ż¼ę“Č°╝Ü░¹ķgėąŽÓ╗źłFŠ█(╬³ę²)Ą─┌ģä▌; Č°ėH╦«╝Ü░¹ätĖ„ūį╩▄ĄĮ╦«ĘųūėĄ─Ī░╬³ę²Ī▒Ż¼ę“Č°╝Ü░¹ķgėąĘų╔óį┌╦«╚▄ę║ųąĄ─┌ģä▌(ŽÓ╗ź┼┼│Ō). įÕ╝Ü░¹ķgĄ─ņoļŖ│Ō┴”ę╗░Ń┤¾ė┌ĘČĄ┬╚Aę²┴”Ż¼ę“┤╦į┌ø]ėą═Ō╝ėą§─²ä®Ž¹│²ņoļŖ│Ō┴”Ą─ŪķørŽ┬Ż¼Lewis╦ß-ēA╦«║Žū„ė├┴”(▒Ē├µėH/╩Ķąį)Ą─ąį┘|║═┤¾ąĪī”╬óįÕæęĖĪę║Ą─Š█╝»ĘĆČ©ąįŠ═Š▀ėąøQČ©ąįĄ─ė░ĒæŻ¼└²╚ń: ėH╦«ąįįÕ╝Ü░¹ķgĄ─Lewis╦ß-ēA╦«║Žū„ė├┴”×ķ│Ō┴”Ż¼ę“┤╦įōŅÉįÕ╝Ü░¹æęĖĪę║┐é╩Ū─▄▒Ż│ųŠ█╝»ĘĆČ©ąį; ų╗ėąLewis╦ß-ēA╦«║Žū„ė├┴”×ķę²┴”Ģr(╩Ķ╦«įÕ╝Ü░¹ų«ķg╝░╠žČ©ėH╦«-╩Ķ╦«įÕ╝Ü░¹ĮM║Ž)Ż¼╬óįÕæęĖĪę║▓┼ėą┐╔─▄░l╔·ą§─². ╝Ü░¹ėH/╩Ķ╦«ąį╚ĪøQė┌Ųõ▒Ē├µ╣”─▄łF: ▒Ē├µĖ╗║¼ķLµ£¤NŅÉĄ─╬óįÕĘNī┘(╚ńŻ¼ŲŽ╠čįÕī┘)▒Ē¼F×ķ╩Ķ╦«ąįŻ¼ę“×ķķLµ£¤NŅÉų„ę¬░³║¼╝ū╗∙║═üå╝ū╗∙Ą╚╩Ķ╦«╗∙łFŻ¼Č°┴u╗∙║═¶╚╗∙Ą╚ėH╦«╗∙łFų╗š╝║▄ąĪę╗▓┐Ęų; ▒Ē├µĖ╗║¼╠Ū╚®╦ßĪóųąąį╠Ū║═ŲŽ╠Ū░ĘĄ╚│╔ĘųĄ─╬óįÕĘNī┘(╚ńąĪŪ“įÕ)ät▒Ē¼F×ķėH╦«ąįŻ¼ę“×ķ▀@ą®│╔Ęų─▄ą╬│╔┤¾┴┐┴u╗∙Īó¶╚╗∙║═░▒╗∙Ą╚ėH╦«╗∙łF.
ĪĪĪĪ2.3 ą§─²ÖC└Ē
ĪĪĪĪĖ∙ō■╔Ž╩÷XDLVO└ĒšōŻ¼╬óįÕą§─²Ą─╗∙▒ŠįŁ└ĒŠ═╩Ūę¬═©▀^ĮĄĄ═/Ž¹│²ņoļŖ│Ō┴”(ZetaļŖ╬╗)Ż¼╩╣Lewis╦ß-ēA╦«║Žū„ė├┴”▒Ē¼F×ķę²┴”Ą╚┤ļ╩®Ž¹│²/ĮĄĄ═įÕ╝Ü░¹ų«ķg▒Ē├µ─▄Ą─┼┼│Ō─▄ĘÕŻ¼╩╣įÕ╝Ü░¹─▄ŽÓ╗ź┐┐Į³ĄĮ▀_Ą┌ę╗Ą═╬╗č©─▄Ż¼Å─Č°Šo├▄Ąžš│ĮYį┌ę╗Ųą╬│╔ą§¾w. ŲõųąŻ¼═Ō╝ė¤oÖCą§─²ä®Ą─ų„ę¬ū„ė├ÖC└ĒŠ═╩Ūųą║═įÕ╝Ü░¹▒Ē├µĄ─ļŖžōąįŻ¼ĮĄĄ═/Ž¹│²ņoļŖ│Ō┴”. ═Ō╝ėĖ▀ĘųūėėąÖCą§─²ä®ätų„ę¬═©▀^╬³ĖĮ╝▄ś“įŁ└ĒŲū„ė├: µ£ĀŅĖ▀Ęųūė╬’┘|(╔┘öĄŪķørę▓┐╔─▄╩Ū¤oÖCą§─²ä®ą╬│╔Ą─┤¾─z┴Ż)į┌ņoļŖę²┴”ĪóĘČĄ┬╚A┴”║═ÜõµI┴”Ą─ū„ė├Ž┬Ż¼ę╗Č╦╬³ĖĮ┴╦─│ę╗─z┴Ż║¾Ż¼┴Ēę╗Č╦ėų╬³ĖĮ┴╦┴Ēę╗─z┴ŻŻ¼Å─Č°░č▓╗═¼Ą──z┴Ż▀BĮėŲüĒČ°ą╬│╔ą§¾w(łD 1). ╔·╬’ą§─²ä®║═EPSšTī¦Ą─ūįą§─²ät┐╔─▄╩Ū═©▀^Lewis╦ß-ēA╦«║Žū„ė├┴”ųąĄ─╩Ķ╦«ę²┴”╝░╬³ĖĮ╝▄ś“įŁ└ĒĄ─ŠC║Žū„ė├īŹ¼Fą§─². ūŅ║¾Ż¼═Č╝ėą§─²ä®ą╬│╔Ą─│┴ĄĒ╬’║═ą§¾wĄ╚▀Ć┐╔═©▀^ŠW▓Č║═ŠĒÆ▀Ą╚╬’└Ēū„ė├▀Mę╗▓Į┤┘▀MįÕ╝Ü░¹Ą─ą§─²│┴ĮĄ.
ĪĪĪĪłD 1 ╬³ĖĮ╝▄ś“ū„ė├╩ŠęŌłD
ĪĪĪĪ3 ═Ō╝ėą§─²ä®Ę© 3.1 ¤oÖCą§─²ä® 3.1.1 ¤oÖCą§─²ä®Ą─ĘNŅÉ║═ū„ė├ÖC└Ē
ĪĪĪĪęįĶF¹}║═õX¹}×ķ┤·▒ĒĄ─ČÓārĮī┘¹}ŅÉ║═Š█║ŽĮī┘¹}ŅÉ╩Ūé„Įy╦«╠Ä└Ēųąæ¬ė├ūŅ×ķÅVĘ║Ą─ą§─²ä®Ż¼ę▓╩Ū╬óįÕą§─²ųąæ¬ė├ūŅįńĄ─═Ō╝ėą§─²ä®. Ąõą═ą§─²ä®░³└©Ż║┴“╦ßõXŻ¼┴“╦ßüåĶFŻ¼┬╚╗»ĶFŻ¼Š█║Ž┬╚╗»õXŻ¼Š█║Ž┴“╦ßõXŻ¼Š█║Ž┴“╦ßõXĶFŻ¼Š█║Ž┬╚╗»õXĶFŻ¼Š█║Ž┴“╦ß┬╚╗»õXĶFĄ╚.
ĪĪĪĪĮī┘¹}ŅÉą§─²ä®ų„ę¬╩Ū═©▀^ļŖąįųą║═ū„ė├ŲŲē─įÕ╝Ü░¹Ą─Š█╝»ĘĆČ©ąį. Al3+ĪóFe3+Ą╚ė╬ļxĻ¢ļxūė╝░ŲõĖ„ĘNĦš²ļŖ║╔Ą─╦«ĮŌ«a╬’─▄ųą║═įÕ╝Ü░¹▒Ē├µ╦∙ĦĄ─žōļŖ║╔Ż¼Å─Č°┤┘▀MįÕ╝Ü░¹┼÷ū▓Š█╝»ą╬│╔ą§¾wŻ¼░l╔·ą§─²│┴ĄĒ.┤╦═ŌŻ¼Al3+Ż¼Fe3+Ą╚Įī┘¹}▀Ć─▄ą╬│╔[Al(OH)3]nĪó[Fe(OH)3]nĄ╚Š█║Ž¾wŻ¼ęį╬³ĖĮ╝▄ś“ą╬╩Įū„ė├ė┌įÕ╝Ü░¹. į┌╠žČ©pHŽ┬Ż¼▀@ą®Įī┘¹}ŅÉ▀Ć┐╔ą╬│╔┤¾┴┐Al(OH)3╝░Fe(OH)3Ą╚│┴ĄĒ╬’Ż¼ęįŠW▓ČŠĒÆ▀ū„ė├┤┘▀M╬óįÕĄ─ą§─²│┴ĮĄ. Š█║ŽĮī┘¹}ŅÉą§─²╬óįÕĄ─ų„ę¬ÖC└Ēät╩Ū╬³ĖĮ╝▄ś“ū„ė├Ż¼═¼Ģrę▓ėąļŖąįųą║═╝░ŠW▓ČŠĒÆ▀ū„ė├.
ĪĪĪĪ3.1.2 ¤oÖCą§─²ä®Ą─ą§─²ą¦╣¹║═ė░Ēæę“╦ž
ĪĪĪĪ▒Ē 1┐éĮY┴╦ÄūĘNĄõą═¤oÖCą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąĄ─ą§─²Śl╝■╝░ą¦╣¹. ė╔ė┌Ė„蹊┐▓╔ė├┴╦▓╗═¼Ą─ėŗ┴┐╗∙£╩(╚ńŻ¼╔·╬’┴┐ęį╝Ü░¹éĆöĄ Īż mL-1╗“mg Īż L-1ėŗ)╝░▓╗═¼Ą─įÕĘN║═ą§─²Śl╝■Ż¼¤oĘ©ų▒Įė▀MąąÖMŽ“▒╚▌^Ż¼Ą½╚į┐╔ęį┐éĮY│÷ę╗ą®ę¬³cŻ║ó┘õX¹}ĪóĶF¹}Ą╚ČÓārĮī┘ą§─²ä®į┌║Ž▀mĄ─Śl╝■Ž┬Č╝┐╔ėąą¦ą§─²(>80%)│ŻęŖĄ─╬óįÕĘNī┘;ī”ė┌Ąõą═Ą─ŽĪįÕę║(ØŌČ╚Ī½0.5 g Īż L-1)Ż¼╦Ää®═Č╝ė┴┐ę╗░Ńę¬▀_Äū░┘mg Īż L-1įÕę║. ó┌ õX¹}▒╚ĶF¹}Ą─ą§─²ą¦┬╩Ė³Ė▀;Įī┘┬╚╗»╬’▒╚Įī┘┴“╦ß¹}Ą─ą§─²ą¦┬╩Ė³Ė▀;▀@Ę┤ė│į┌▀_ĄĮŅÉ╦Ųą§─²ą¦╣¹Ż¼õX¹}║═Įī┘┬╚╗»╬’Ą─═Č╝ė┴┐Ė³ąĪŪę╦∙ąĶą§─²ĢrķgĖ³Č╠. ó█Š█║ŽĮī┘¹}ŅÉ▒╚ĘŪŠ█║ŽĮī┘¹}ŅÉĄ─╗ņ─²ą¦┬╩Ė▀Ż¼Ūęį┌Ė³ÅVĄ─pHųĄĘČć·ā╚ėąą¦.
| ą§─²ä® | įÕę║ | ą§─²Śl╝■ | ą§─²ą¦╣¹Ż║╚ź│²┬╩(═Č╝ė┴┐)Ż¼ą§─²Ģrķg | ģó┐╝╬─½I |
| å╬╬╗Ż║1. å╬╬╗×ķmmol Īż L -1 (ęįAl╗“Feėŗ)Ż╗ 2. å╬╬╗×ķg Īż L -1 . | ||||
| ┴“╦ßõX ┴“╦ßĶF Š█║Ž┬╚╗»õX Š█║Ž┴“╦ßĶF |
ĒŚ╚”įÕ( Anabaena )Ż¼ įÕę║ØŌČ╚2Ī┴10 6 cells Īż mL -1 |
2 min┐ņ╦┘öć░Ķ(300 r Īż min -1 )Ż¼ 25 min┬²╦┘öć░Ķ(35 r Īż min -1 )Ż¼ 2 h │┴ĄĒŻ¼pH 7.5; ═Č╝ė┴┐ 1 Ż║┴“╦ßõX(0.175Ż¼0.25Ż¼0.375)Ż¼┴“╦ßĶF(0.175Ż¼0.21Ż¼0.25)Ż¼Š█║Ž┬╚╗»õX(0.175Ż¼0.26Ż¼0.375)Ż¼Š█║Ž┴“╦ßõX(0.175Ż¼0.21Ż¼0.25) |
┴“╦ßõXŻ║ 74%(0.175)Ż¼94%(0.25)Ż¼95%(0.375); ┴“╦ßĶFŻ║ 70%(0.175)Ż¼75%(0.21)Ż¼76%(0.25); Š█║Ž┬╚╗»õXŻ║ 67%(0.175)Ż¼69%(0.26)Ż¼73%(0.375); Š█║Ž┴“╦ßĶFŻ║ 94%(0.175)Ż¼95%(0.21)Ż¼96%(0.25) |
Jiang et al.Ż¼ 1993 |
| ┴“╦ßõX ┴“╦ßĶF ┬╚╗»õX ┬╚╗»ĶF |
╬óąĪąĪŪ“įÕ( Chlorella minutissima )Ż¼įÕę║ØŌČ╚220Ī┴10 6 cells Īż mL -1 |
═Č╦Ä┴┐ 2 Ż║0.25Ż¼0.50Ż¼0.75Ż¼1 | ┴“╦ßõXŻ║80%(0.75)Ż¼2 h; ┴“╦ßĶFŻ║80%(0.75)Ż¼4 h; ┬╚╗»õXŻ║80%(0.5)Ż¼1 h; ┬╚╗»ĶFŻ║80%(0.5)Ż¼3 h; |
Papazi et al.Ż¼ 2010 |
| ┴“╦ßõX ┴“╦ßĶF ╚²┬╚╗»ĶF Üõč§╗»Ō} |
ąĪŪ“įÕ( Chlorella )Ż¼ įÕę║ØŌČ╚0.53 g Īż L -1 |
═Č╝ė┴┐ 2 Ż║┴“╦ßõX(0.8)Ż¼┴“╦ßĶF(0.5)Ż¼ ╚²┬╚╗»ĶF(0.3)Ż¼Üõč§╗»Ō}(0.8) |
┴“╦ßõXŻ║89.7%(0.8)Ż¼90 min; ┴“╦ßĶFŻ║89.6%(0.5)Ż¼90 min; ╚²┬╚╗»ĶFŻ║92.3%(0.3)Ż¼30 min; Üõč§╗»Ō}Ż║91.7%(0.8)Ż¼90 min |
č”╚žĄ╚Ż¼ 2012 |
| ┴“╦ßõX ┬╚╗»ĶF Üõč§╗»Ō} |
¢┼įÕ( Scenedesmus sp.)Ż¼ įÕę║ØŌČ╚0.54 g Īż L -1 |
1 min┐ņ╦┘öć░Ķ(800 r Īż min -1 )Ż¼1 min┬²╦┘öć░Ķ(250 r Īż min -1 )Ż¼│┴ĄĒĢrķg(2Ż¼5Ż¼10Ż¼30Ż¼60Ż¼120 min)Ż╗═Č╝ė┴┐ 2 Ż║┴“╦ßõX(0.02Ż¼0.03Ż¼0.05Ż¼0.1Ż¼0.3)Ż¼┬╚╗»ĶF(0.06Ż¼0.08Ż¼0.1Ż¼0.15Ż¼0.2)Ż¼Üõč§╗»Ō}(0.2Ż¼0.3Ż¼0.4Ż¼0.5Ż¼0.6) | ┴“╦ßõXŻ║ >95%(0.3)Ż¼10 min; 75%(0.1)Ż¼30 min; Ī½60%(0.02Ż¼0.03Ż¼0.05)Ż¼120 min ┬╚╗»ĶFŻ║ >95%(0.15Ż¼2)Ż¼2 min; Ī½70%(0.06Ż¼0.08Ż¼0.1)Ż¼120 min Üõč§╗»Ō}Ż║ 90%(0.3Ż¼0.4)Ż¼120 min; Ī½80%(0.5)Ż¼120 min; Ī½60%(0.6)Ż¼120 min | |
ĪĪĪĪ▒Ē 1 ¤oÖCą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąĄ─ą§─²Śl╝■┼cą¦╣¹
ĪĪĪĪė░Ēæ¤oÖCą§─²ä®ą§─²ą¦┬╩Ą─ę“╦žų„ę¬ėąŻ║ą§─²ä®Ą─ĘNŅÉĪópHĪóįÕę║ØŌČ╚║══Č╝ė┴┐Ą╚. ą§─²ä®ĘNŅÉī”ą§─²ą¦╣¹Ą─ė░Ēæį┌╔Ž╬─ęčėą╦∙šō╩÷. ę╗░ŃČ°čįą§─²ä®╦∙ĦĄ─ļŖ║╔├▄Č╚įĮĖ▀Ż¼ą§─²ą¦╣¹įĮ║├;▀@š²╩ŪõX¹}ĪóĶF¹}Ą─ą§─²ą¦╣¹ę¬▀h▀h║├ė┌Ō}ĪóµV║═õ@ļxūėĄ─įŁę“╦∙į┌. ┤╦═ŌŻ¼ą§─²ä®Ą─╦«╚▄ąįę▓ī”Ųõą§─²ą¦╣¹ėą’@ų°ė░ĒæŻ¼╚ńŻ║┬╚ļxūėĄ─╦«╚▄ąį║├ė┌┴“╦ßĖ∙Ż¼╦∙ęįĮī┘┬╚╗»╬’Ą─ą§─²ą¦╣¹ę¬║├ė┌Įī┘┴“╦ß¹}.╦«╚▄ąįą¦æ¬▀Ć┐╔ęįĮŌßī─”Ā¢┘|┴┐ī”ą§─²ą¦╣¹Ą─ė░ĒæŻ¼╚ńŻ║▒M╣▄Fe3+║═Al3+ ę╗śėĦš²3ārļŖ║╔Ż¼Ą½ĶFļxūėĄ──”Ā¢┘|┴┐┤¾ė┌õXļxūė;─”Ā¢┘|┴┐įĮ┤¾Ż¼╦«╚▄ąįįĮ▓Ņ;╦∙ęįĶF¹}Ą─ą§─²ą¦┬╩ꬥ═ė┌õX¹}. õX¹}Ą─ą§─²ą¦╣¹ī”pHĖ▀Č╚├¶ĖąŻ¼ūŅ╝čpH×ķ4~5Ż¼▀@╩Ūę“×ķį┌┤╦pHŚl╝■Ž┬õXĄ─╦«ĮŌ«a╬’ęįĦš²ļŖĄ─ČÓ║╦┴u╗∙┼õ║Ž╬’ą╬╩Į┤µį┌ŪęūŅĘĆČ©;ųąąįŚl╝■Ž┬Ż¼õXĄ─╦«ĮŌ«a╬’ęįAl(OH)3│┴ĄĒ×ķų„;pH>8.5ĢrŻ¼╦«ĮŌ«a╬’īóęįĦžōļŖĄ─[Al(OH)4]-×ķų„Ż¼¤oĘ©ą╬│╔ėąą¦ą§─². ę╗Č©ĘČć·ā╚ą§─²ą¦╣¹┼cą§─²ä®Ą─═Č╝ė┴┐│╔š²▒╚; Ą½▀^┴┐═Č╝ėĢ■╩╣─z┴Ż╬³ĖĮ▀^ČÓĄ─Ę┤ļxūėŻ¼ųžą┬ĦļŖČ°į┘┤╬ĘĆČ©; ę“┤╦ą§─²ä®╩╣ė├┤µį┌ę╗éĆūŅ╝č═Č╝ė┴┐Ż¼▀@╩Ūą§─²ĘųļxųąįńęčĄ├ĄĮ╔Ņ╚ļĘų╬÷Ą─Ąõą═¼FŽ¾. Ą½ūŅ╝č═Č╝ė┴┐ļSįÕę║ØŌČ╚Ą─ūā╗»ęÄ┬╔╬─½Iųą▀Ć┤µį┌▓╗Įyę╗ų«╠ÄŻ║ę╗░ŃČ°čį╦∙ąĶą§─²ä®Ą─═Č╝ė┴┐ļSįÕę║ØŌČ╚Ą─╔²Ė▀Č°ŠĆąįį÷╝ėŻ¼▀@Ę¹║ŽļŖąįųą║═Ą─ą§─²ÖC└Ē. ╚╗Č°Ż¼Garzon-SanabriaĄ╚į┌╩╣ė├┬╚╗»õXą§─²╬óŠGŪ“įÕĢr░l¼FŻ¼«öįÕę║ØŌČ╚║▄Ė▀ĢrŻ¼▀_ĄĮ═¼śėą§─²ą¦╣¹Ą─═Č╝ė┴┐ģsöĄ▒ČĄžĄ═ė┌ŽĪįÕę║. WyattĄ╚į┌ė├┬╚╗»ĶFą§─²ąĪŪ“įÕĢrę▓Ą├│÷┴╦ŅÉ╦ŲĄ─ĮYšō. ŲõįŁę“║▄┐╔─▄╩Ūį┌Ė▀įÕę║ØŌČ╚Ģrą╬│╔┴╦’@ų°Ą─ŠW▓Č║═ŠĒÆ▀ū„ė├. ūŅ║¾Ż¼ūŅą┬蹊┐▒Ē├„╬óįÕ┤·ųx«a╔·Ą─ėąÖC╬’ī”ą§─²▀^│╠ėą’@ų°Ą─ęųųŲū„ė├Ż¼Ųõ┤µį┌īó│╔▒ČĄžį÷╝ėą§─²ä®═Č╝ė┴┐Ż¼▀@īó’@ų°į÷╝ėą§─²│╔▒Š▓óī”įÕ╝Ü░¹Ą─║¾└m╝ė╣ż└¹ė├įņ│╔žō├µė░Ēæ.
ĪĪĪĪ3.2 ėąÖCĖ▀Ęųūėą§─²ä®Ę© 3.2.1 ĘNŅÉ┼cū„ė├ÖC└Ē
ĪĪĪĪėąÖCĖ▀Ęųūėą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąęÓ║▄įń▒ŃĄ├ĄĮ┴╦æ¬ė├. ─┐Ū░╔╠śI╗»Ą─ėąÖCĖ▀Ęųą§─²ä®ų„ę¬×ķ╚╦╣ż║Ž│╔Ż¼ęįŠ█▒¹Ž®§Ż░Ę(Polyacrylamide)×ķ┤·▒Ē. Į³─ĻüĒŻ¼╠ņ╚╗Ė▀ĘųūėėąÖCą§─²ä®Ż¼╚ńŻ¼ÜżŠ█╠Ū(Chitosan)ĪóĻ¢ļxūėĄĒĘ█(Cationic Starch)║═└wŠS╦žĄ╚Ą├ĄĮ┴╦įĮüĒįĮČÓĄ─ĻPūó. ėąÖCĖ▀Ęųūėą§─²ä®Ą─ū„ė├ÖC└Ēų„ę¬×ķ╬³ĖĮ╝▄ś“ū„ė├. ę“įÕ╝Ü░¹Ä¦žōļŖĄ─▒Ē├µ╠žąįŻ¼Ė▀ą¦Ą─Ė▀Ęųūėą§─²ä®▒žĒÜ×ķĻ¢ļxūėą═Ą─. ĻÄļxūė╝░ĘŪļxūėą═Ą─Š█║ŽĖ▀Ęųūėå╬¬Ü╩╣ė├Ģr▓╗─▄╩╣╬óįÕ░l╔·ėąą¦ą§─². │²╝▄ś“ū„ė├═ŌŻ¼Ļ¢ļxūėą═Ė▀Ęųūėą§─²ä®▀Ć┐╔─▄Šų▓┐─µ▐DįÕ╝Ü░¹▒Ē├µĄ─ļŖžōąįŻ¼╩╣Ųõ─│ą®▓┐╬╗ĦžōļŖČ°┴Ēę╗▓┐╬╗Ħš²ļŖ;Å─Č°╩╣▓╗═¼Ą─įÕ╝Ü░¹─▄ų▒Įė═©▀^ņoļŖę²┴”ĮY║Žį┌ę╗ŲŻ¼ą╬│╔╦∙ų^Ą─ņoļŖ╗źčaŠ█╝». ęįŽ┬ĮķĮBÄūĘN┤·▒ĒąįĄ─ėąÖCĖ▀Ęųūėą§─²ä®.
ĪĪĪĪa. Š█▒¹Ž®§Ż░Ę
ĪĪĪĪŠ█▒¹Ž®§Ż░ĘĘųūė┴┐į┌400~2000╚fų«ķgŻ¼Š▀ėąĻ¢ąį╗∙łF(Ī¬CONH2). įō╗∙łF╝╚╩ŪėH╦«╗∙łFŻ¼ėų╩Ū╬³ĖĮ╗∙łFŻ¼╦∙ęį─▄ī”╬óįÕ«a╔·╬³ĖĮļŖųą║═╝░╝▄ś“ū„ė├. │²ś“▀Bū„ė├═ŌŻ¼Š█▒¹Ž®§Ż░Ę▀Ćėą░³Įjū„ė├. ░l╔·ś“▀B║═░³ĮjĄ─Ė▀Ęųūė─▄ą╬│╔╚²ŠSŠWĀŅĮYśŗŻ¼═©▀^ŠĒÆ▀ŠW▓Čū„ė├╩╣╬óįÕ│┴ĮĄĘųļx.
ĪĪĪĪb. ÜżŠ█╠Ū
ĪĪĪĪÜżŠ█╠Ū╩Ūī”╝ūÜż╦ž▀Mąą├ōęꧯ╗∙Č°Ą├ĄĮĄ─Ż¼╩Ū╔┘öĄĻ¢ļxūėą═Ą─╠ņ╚╗Ė▀Š█╬’. ŲõĮYśŗå╬į¬╩Ū2-░▒╗∙-2├ōč§ŲŽ╠č╠ŪŻ¼═©▀^”┬-1-4╠Ū▄šµI▀BĮėŲüĒ(łD 2). į┌╦ßąįŚl╝■Ž┬Ż¼ÜżŠ█╠ŪĘųūėµ£╔Ž╦∙ĦĄ─┤¾┴┐░▒╗∙ęįĦš²ļŖ║╔Ą─░Ęļxūėą╬╩Į┤µį┌Ż¼─▄ųą║═įÕ╝Ü░¹Ą─ļŖžōąįŻ¼═¼ĢrĮĶų·Ė▀Ęųūėµ£Ą─╬³ĖĮ╝▄ś“ū„ė├╩╣įÕ¾wą§─²│┴ĮĄ. «ö╚▄ę║│╩¼FēAąįĢrŻ¼ÜżŠ█╠Ū▒Ē├µ╦∙Ħ░Ę╗∙ĘŪļxūė╗»╗“│╩╚§žōļŖąįŻ¼Å─Č°ĮĄĄ═┴╦Ųõą§─²ą¦┬╩(└Ņ╚¶╗█Ą╚Ż¼2012).
ĪĪĪĪłD 2 ÜżŠ█╠ŪĮYśŗ
ĪĪĪĪc. Ļ¢ļxūėĄĒĘ█
ĪĪĪĪĻ¢ļxūėĄĒĘ█╩Ūį┌ĄĒĘ█╣Ū╝▄ųąę²╚ļ╝Šõ@╗∙łFŻ¼▀@śėŠ═╩╣Ą├ĄĒĘ█│╩š²ļŖąį. ėųę“ĄĒĘ█Ęųūė╣╠ėąĄ─Š█║ŽĮYśŗŻ¼╩╣Ļ¢ļxūėĄĒĘ█Š▀ėąļŖąįųą║═╝░╬³ĖĮ╝▄ś“Ą─ļpųžū„ė├. Ļ¢ļxūėĄĒĘ█║═ÜżŠ█╠Ūę╗śėŻ¼ę▓Š▀ėą¤oČŠĪó¤o╬█╚ŠĪó┐╔╔·╬’ĮĄĮŌĄ─╠ž³c. ┼cÜżŠ█╠Ū▒╚▌^Č°čįŻ¼Ļ¢ļxūėĄĒĘ█įŁ┴ŽārĖ±Ė³Ą═Ż¼Ė³╚▌ęū½@Ą├. ūŅ×ķ’@ų°Ą─╩ŪŲõ╝Š░Ę╗∙łF▓╗╩▄pHĄ─ė░ĒæŻ¼Å─Č°╩╣Ųõ┐╔į┌║▄īÆĄ─pHųĄĘČć·ā╚▀mė├.
ĪĪĪĪ3.2.2 ą§─²ą¦╣¹┼cė░Ēæę“╦ž
▒Ē 2┴ą│÷┴╦ÄūĘNĄõą═ėąÖCĖ▀Ęųūėą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąĄ─ą§─²Śl╝■┼cą¦╣¹. ▒Ē 2’@╩ŠŻ¼Š█▒¹Ž®§Ż░Ęļm╚╗╩Ū╦«╠Ä└Ēųąæ¬ė├ūŅ│╔╩ņĄ─Ė▀Ęųūėą§─²ä®Ż¼Ą½Ųõī”╬óįÕĄ─ą§─²ą¦┬╩ģs▓ó▓╗└ĒŽļ; į┌ŽÓī”▌^Ė▀Ą─═Č╝ė┴┐Ž┬(20~80 mg Īż L-1įÕę║)ę▓āH─▄īŹ¼F50%ū¾ėęĄ─ą§─²ą¦╣¹. ▀@┐╔─▄╩Ūę“×ķŲõļŖ║╔├▄Č╚▌^Ą═╦∙ų┬. ę“┤╦ī”Š█▒¹Ž®§Ż░Ę▀MąąĖ─ąį╩Ūę╗éĆųžę¬Ą─蹊┐ĘĮŽ“Ż¼╝┤Ż¼═©▀^į┌Š█▒¹Ž®§Ż░Ę╔Žę²╚ļ░ĘŅÉĘųūėŻ¼╔·│╔╝Š░Ęą═Ļ¢ļxūėęį▀Mę╗▓Į╠ßĖ▀ą§─²ą¦┬╩║═▀mė├ĘČć·. ┤╦═ŌŻ¼ChenĄ╚šJ×ķŠ█▒¹Ž®§Ż░ĘĄ─Ė▀Ęųūėµ£š╣ķ_│╠Č╚▓╗ūŃ╩Ūī¦ų┬Ųõą§─²ą¦╣¹▓╗╝čĄ─įŁę“.ę“┤╦Ż¼Å─ĮYśŗ╔Žī”Š█▒¹Ž®§Ż░Ę▀MąąĖ─▀MŻ¼į÷ÅŖĖ▀Ęųūėµ£Ą─š╣ķ_│╠Č╚ę▓╩ŪÅŖ╗»Ųõą§─²ą¦╣¹Ą─ĘĮŽ“ų«ę╗.
| ą§─²ä® | įÕę║ | ą§─²Śl╝■ | ą§─²ą¦╣¹ | ģó┐╝╬─½I |
| Š█▒¹Ž®§Ż░Ę | ¢┼įÕ( Scenedesmus sp.)Ż¼įÕę║ØŌČ╚0.54 g Īż L -1 |
═Č╝ė┴┐: 20Ī½80 mg Īż L -1 ; ą§─²Ģrķg120 min |
Ė„═Č╝ė┴┐Ą─ą§─²ą¦╣¹╗∙▒Šę╗ų┬Ż¼╝s50% | Chen et al.Ż¼ 2013 |
| ÜżŠ█╠Ū | ╬ó─ęįÕ( Microcystis )Ż¼įÕę║ØŌČ╚5Ī┴10 5 Ī½2Ī┴10 6 cells Īż mL -1 | ═Č╝ė┴┐: 0.1Ī½1 mg Īż L -1 ; pH 4Ī½9; 1 min┐ņ╦┘öć░Ķ(300 r Īż min -1 )Ż¼10 minųą╦┘öć░Ķ(100 r Īż min -1 )Ż¼10 min┬²╦┘öć░Ķ(50 r Īż min -1 )Ż¼│┴ĄĒ30 min | ═Č╝ė┴┐>0.5 mg Īż L -1 Ģrą§─²ą¦╣¹>90%; ▀mę╦pH 5Ī½7Ż¼ūŅ╝č×ķ6Ż¼┤¾ė┌8Ģr╗∙▒Š¤oą§─²ą¦╣¹ | Ąį½hĄ╚Ż¼ 2009 |
| ÜżŠ█╠Ū | ąĪŪ“įÕ( Chlorella sp.)Ż¼įÕę║ØŌČ╚5Ī┴10 9 cells Īż mL -1 |
═Č╝ė┴┐: 5Ī½100 mg Īż L -1 ; öć░Ķ60 min(100 r Īż min -1 )Ż¼│┴ĄĒ60 min | Ą══Č╝ė┴┐Ģrą§─²ą¦╣¹ļS═Č╝ė┴┐ŠĆąį╔Ž╔²Ż¼ūŅ╝č═Č╝ė┴┐×ķ10 mg Īż L -1 (99%╚ź│²)Ż¼└^└m╝ė┤¾═Č╝ė┴┐ą§─²ą¦╣¹Ž┬ĮĄ | Ahmad et al.Ż¼ 2011 |
| Ļ¢ļxūėĄĒĘ█ (Greenfloc 120) |
öMąĪŪ“įÕī┘ ( ParaChlorella )Ż¼ ą▒╔·¢┼įÕ( Scenedesmus obliquus ); įÕę║ØŌČ╚0.075Ī½ 0.3 g Īż L -1 |
═Č╝ė┴┐: 5Ī½60 mg Īż L -1 ; pH 5Ī½10; 5 min┐ņ╦┘öć░Ķ(1000 r Īż min -1 )Ż¼ 25 minųą╦┘öć░Ķ(250 r Īż min -1 )Ż¼ │┴ĄĒ30 min |
öMąĪŪ“įÕī┘Ż║ą§─²ą¦╣¹▀_80%ęį╔ŽĄ─╦Ää®═Č╝ė┴┐┼c╔·╬’┴┐ų«▒╚╝s×ķ0.1(įÕę║ØŌČ╚0.3 g Īż L -1 ī”æ¬═Č╝ė┴┐30 mg Īż L -1 ); ▀^┴┐═Č╝ėįņ│╔├„’@Ą──z¾w▒Żūoū„ė├; ą§─²ą¦╣¹╗∙▒Š▓╗╩▄pHė░Ēæ. ą▒╔·¢┼įÕŻ║ą§─²ą¦╣¹▀_80%ęį╔ŽĄ─╦Ää®═Č╝ė┴┐┼c╔·╬’┴┐ų«▒╚×ķ0.03(įÕę║ØŌČ╚0.3 g Īż L -1 ī”æ¬═Č╝ė┴┐9 mg Īż L -1 ) | V and amme et al .Ż¼2010 |
| Ļ¢ļxūėĄĒĘ█ (ė±├ūĄĒĘ█) |
ą▒╔·¢┼įÕ( Scenedesmus obliquus )Ż¼įÕę║ØŌČ╚0.2Ī½0.25 g Īż L -1 | ═Č╝ė┴┐/╬óįÕ╔·╬’┴┐Ż║0Ī½0.18; pH 7; 2 min┐ņ╦┘öć░Ķ(200 r Īż min -1 )Ż¼10 min┬²╦┘öć░Ķ(25 r Īż min -1 )Ż¼│┴ĄĒ1 h | į┌═Č╝ė┴┐/╬óįÕ╔·╬’┴┐ų«▒╚×ķ0.0053ĢrŠ═▀_ĄĮ┴╦90%Ą─ą§─²ą¦╣¹(įÕę║ØŌČ╚0.25 g Īż L -1 ī”æ¬═Č╝ė┴┐1.3 mg Īż L -1 ) | Anthony et al .Ż¼2013 |
ĪĪĪĪ▒Ē 2 ėąÖCĖ▀Ęųūėą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąĄ─ą§─²Śl╝■╝░ą¦╣¹
ĪĪĪĪÜżŠ█╠Ū║═Ļ¢ļxūėĄĒĘ█ī”ĄŁ╦«įÕŅÉČ╝ėąĘŪ│ŻĖ▀Ą─ą§─²ą¦┬╩: ę╗░Ńį┌10~30 mg Īż L-1įÕę║Ą─═Č╝ė┴┐Ž┬Š═┐╔ęį▀_ĄĮ80%ęį╔ŽĄ─ą§─²ą¦╣¹; ī”ė┌éĆäeįÕĘN╔§ų┴į┌1 mg Īż L-1ū¾ėęĄ─═Č╝ė┴┐Ž┬Š═─▄▀_ĄĮ90%ęį╔ŽĄ─ą§─²ą¦╣¹. ▀@▒╚▀_ĄĮ═¼śėą§─²ą¦╣¹Ą─¤oÖCą§─²ä®═Č╝ė┴┐(▒Ē 1)ꬥ═ę╗éĆöĄ┴┐╝ēęį╔Ž. ŲõįŁę“ų„ę¬╩ŪĖ▀Ęųūėą§─²ä®Š▀ėą’@ų°Ą─╬³ĖĮ╝▄ś“ū„ė├Ż¼ę“┤╦┐╔ęįį┌įÕ╝Ü░¹žōļŖąį▀h╬┤▒╗ųą║═Ą─ŪķørŽ┬(ZetaļŖ╬╗<<0)Š═īŹ¼FĖ▀ą¦ą§─².
ĪĪĪĪė░ĒæĖ▀Ęųūėą§─²ä®ą§─²ą¦╣¹Ą─ų„ę¬ę“╦žėąŻ║─”Ā¢┘|┴┐ĪóļŖ║╔├▄Č╚Īó═Č╝ė┴┐ĪóįÕ╝Ü░¹ØŌČ╚ĪóļxūėÅŖČ╚/¹}Č╚ĪópH║═öć░ĶÅŖČ╚Ą╚. ─”Ā¢┘|┴┐▌^Ė▀Ą─Ė▀Ęųūėą§─²ä®Š▀ėąĖ³ČÓĄ─╬³ĖĮ╝▄ś“ĮY║Ž³cŻ¼ę“┤╦ę╗░ŃŠ▀ėąĖ³║├Ą─ą§─²ą¦╣¹. ļŖ║╔├▄Č╚Ė▀Ą─Ė▀Ęųūėą§─²ä®Š▀ėąĖ³ÅŖĄ─ļŖąįųą║═─▄┴”; ┤╦═ŌŻ¼Ė▀Ą─ļŖ║╔├▄Č╚▀Ćėąų·ė┌Ė▀Ęųūėµ£Ą─│õĘųš╣ķ_Ż¼į÷ÅŖ╝▄ś“─▄┴”. ═Č╝ė┴┐▓╗ūŃŻ¼ą§─²ą¦╣¹▓╗│õĘų; Ą½═Č╝ė▀^┴┐Ż¼ėųĢ■ī”─z┴ŻŲĄĮĘĆČ©▒Żūoū„ė├; ▀@┼c¤oÖCą§─²ä®ųąūŅ╝č═Č╝ė┴┐Ą─Ė┼─Ņę╗ų┬. Ė▀Ą─įÕ╝Ü░¹ØŌČ╚╩╣Ņw┴ŻķgĄ─┼÷ū▓Ė³╝ėŅlĘ▒Ż¼į┌ę╗Č©ĘČć·ā╚īó┤┘▀Mą§─²ū„ė├. ┼c¤oÖCą§─²ä®ŽÓ▒╚Ż¼Ļ¢ļxūėą═Ė▀Š█╬’Ą─ę╗éĆ╠ž³c╩ŪŻ¼Ė▀Ą─ļxūėÅŖČ╚/¹}Č╚ī”Ųõą§─²ą¦╣¹ėą’@ų°Ą─ęųųŲū„ė├.▀@╩Ūę“×ķį┌Ė▀ļxūėÅŖČ╚/¹}Č╚ŪķørŽ┬Ż¼Ļ¢ļxūėą═Ė▀Š█╬’ėąłFŠ█į┌ę╗ŲĄ─┌ģä▌Ż¼╝▄ś“ū„ė├īó’@ų°£p╚§ ▀@╩╣Ųõį┌▓╔╩š║Żč¾╬óįÕĢr╩▄ĄĮŽ▐ųŲ. ÜżŠ█╠Ūę╗░Ńį┌╦ßąįŚl╝■Ž┬ą§─²ą¦╣¹▓┼’@ų°Ż¼▀@═∙═∙│¼│÷┴╦╬óįÕ┼ÓB¾wŽĄĄ─š²│ŻpHĘČć·Ż¼Å─Č°Ž▐ųŲ┴╦Ųõæ¬ė├. Č°Ļ¢ļxūėĄĒĘ█╗∙▒Š▓╗╩▄pHė░ĒæŻ¼į┌pH 5~10Ą─ĘČć·ā╚Č╝─▄ŠS│ų+15 mVū¾ėęĄ─ZetaļŖ╬╗Ż¼Š▀ėąŲš▒ķĄ─▀mė├ąį. Ą═╦┘öć░Ķī”ą╬│╔┤¾Ą─ą§¾wėą└¹; ▀^ÅŖĄ─öć░ĶīóŲŲē─ęčą╬│╔Ą─ą§¾w.
ĪĪĪĪ3.3 ╔·╬’ą§─²ä®Ę©
ĪĪĪĪ╔·╬’ą§─²ä®(Bioflocculant)╩ŪĮ³Äū─Ļ╬óįÕą§─²Ą─蹊┐¤ß³cų«ę╗. ╔·╬’ą§─²ä®ę╗░Ń╩ŪųĖ╬ó╔·╬’┤·ųx╗Ņäėųą«a╔·Ą─Š▀ėąą§─²ą¦╣¹Ą─░¹═ŌŠ█║Ž╬’(EPS). ╝ÜŠ·Ī󚵊·║═Ę┼ŠĆŠ·Č╝╩Ū─▄«a╔·╔·╬’ą§─²ä®Ą─│ŻęŖ╬ó╔·╬’(Lam and LeeŻ¼ 2012). ╔·╬’ą§─²ä®į┌╬óįÕ▓╔╩šųąĄ─Š▀¾wæ¬ė├ĘĮ╩Įų„ę¬░³└©ęįŽ┬ÄūĘNŻ║ó┘═Č╝ėą§─²╬ó╔·╬’Ą─╗ņ║Ž┼ÓBę║(╬ó╔·╬’╝Ü░¹+┼ÓBę║);ó┌Š·-įÕ╗ņ║Ž┼ÓB(ąĶį┌╬óįÕ┼ÓBŽĄĮy╠Ē╝ėėąÖC╠╝į┤);ó█ą§─²╬ó╔·╬’Ą─░¹═Ō│ķ╚Īę║(ļxą─║¾Ą─╔ŽŪÕę║)ū„×ķą§─²ä®;ó▄Ęųļx╝ā╗»║¾Ą─░¹═Ō╠ß╚Ī╬’ū„×ķą§─²ä®;ó▌ų▒Įė═Č╝ėą§─²╬ó╔·╬’╝Ü░¹ū„×ķą§─²ä®.
ĪĪĪĪ▒Ē 3┐éĮY┴╦Ė„ĘN╔·╬’ą§─²ä®į┌╬óįÕą§─²ĘųļxųąĄ─æ¬ė├Ūķør. ▒Ē 3’@╩ŠŻ¼╔·╬’ą§─²ä®Ą─ą§─²ą¦┬╩ęÓ║▄Ė▀Ż¼ę╗░Ńį┌═Č╝ė┴┐×ķ10~30 mg Īż L-1įÕę║Ģr▒Ń┐╔▀_ĄĮ>80%Ą─ą§─²ą¦╣¹Ż¼ū„ė├├„’@║├ė┌¤oÖCą§─²ä®(▒Ē 1). Ą½į┌ą§─²ą¦╣¹Ą─ė░Ēæę“╦ž╔Žģs┤µį┌▓╗╔┘ŽÓ╗ź├¼Č▄ų«╠Ä. ę╗ą®čąŠ┐’@╩Šą§─²ą¦╣¹Ģ■ļSpH╔²Ė▀├„’@╝ėÅŖŻ¼Ą½ę▓ėąčąŠ┐’@╩Šą§─²ą¦╣¹╗∙▒Š▓╗╩▄pHė░Ēæ;ČÓöĄčąŠ┐▒Ē├„Ż¼ČÓārĻ¢ļxūė─▄’@ų°┤┘▀Mą§─²╔§ų┴╩Ūą╬│╔ą§─²Ą─▒žę¬Śl╝■Ż¼Ą½į┌╔┘öĄčąŠ┐ųąČÓārĻ¢ļxūėī”ą§─²ą¦╣¹╗∙▒Šø]ė░Ēæ;LeeĄ╚’@╩Šą§─²╬ó╔·╬’└¹ė├▓╗═¼╠╝į┤«a╔·Ą─ą§─²ä®ą§─²ą¦╣¹╗∙▒Šę╗ų┬Ż¼Č°WangĄ╚ęį▓╗═¼╠╝į┤×ķĄū╬’«a╔·Ą─╔·╬’ą§─²ä®ą§─²ą¦╣¹Õ─«É.▀@ą®├¼Č▄┐╔─▄╩ŪĖ„ĘN╔·╬’ą§─²ä®į┌ĘNŅÉĪóĮM│╔╝░ą§─²ÖC└Ē╔ŽĄ─▓╗═¼Č°ī¦ų┬Ą─.
| üĒį┤ | æ¬ė├ĘĮ╩Į | įÕę║ | ą§─²Śl╝■ | ą§─²ą¦╣¹╝░ė░Ēæę“╦ž | ģó┐╝╬─½I |
| č┐µ▀ŚUŠ· ( Paenibacillus sp.) |
ų▒Įė═Č╝ė┼ÓBę║(įŁę║) | ąĪŪ“įÕ( Chlorella vulgaris )Ż¼įÕę║ØŌČ╚0.062 g Īż L -1 | ═Č╝ė┴┐20 mL Īż L -1 Ż¼Ļ¢ļxūė6.8 mmol Īż L -1 (CaCl 2 Ż¼MgCl 2 Ż¼FeCl 3 Ż¼CaCl 2 Ż¼KClŻ¼NaCl); pH 5Ī½11 | 77%Ī½86%; ą§─²ą¦╣¹ļSpH╔²Ė▀Č°į÷ÅŖ; ČÓārĻ¢ļxūėĄ─ų·─²ą¦╣¹’@ų°║├ė┌å╬ārĻ¢ļxūėŻ¼CaCl 2 ūŅ╝č | Oh et al ., 2001 |
| ╩®╩Ž╝┘å╬░¹Š· ( Pseudomonas stutzeri ) ║═Ž×śėč┐µ▀ŚUŠ· ( Bacillus cereus ) |
Š·-įÕ╗ņ║Ž┼ÓB | Ņw╩»įÕ( Pleurochrysis carterae )Ż¼įÕę║ØŌČ╚ Ī½0.5 g Īż L -1 |
0.1 mLŠ·ę║ + 100 mLįÕę║; ═Ō╝ė╠╝į┤: ęę╦߯¼ŲŽ╠č╠ŪŻ¼Ė╩ė═(0.1 g Īż L -1 ); ą§─²(╣▓═¼┼ÓB)Ģrķg: 6Ż¼24 h | 6 h: 45%Ī½53%; 24 h: 88%Ī½94%; ═Ō╝ė╠╝į┤ĘNŅÉī”ą§─²ą¦╣¹ø]’@ų°ė░Ēæ | Lee et al ., 2009 |
| ČÓš│ŅÉč┐µ▀ŚUŠ· ( Paenibacillus polymyxa ) |
═Č╝ė│ķ╚Īę║(įŁ┼ÓBę║ŽĪßī10▒Č▓óļxą─║¾Ą─╔ŽŪÕę║) | ¢┼įÕ( Scenedesmus sp.)Ż¼įÕę║ØŌČ╚2.35 g Īż L -1 | ═Č╝ė┴┐1%( V/V )+ Ļ¢ļxūė(å╬¬Ü╗“ĮM║Ž)Ż║CaCl 2 Ż¼MgSO 4 Ż¼FeCl 3 Ż¼Al 2 (SO 4 ) 3 | Ļ¢ļxūėå╬¬Ü═Č╝ė: 0.5 mmol Īż L -1 FeCl 3 (35%)>10 mmol Īż L -1 CaCl 2 (18%); Ļ¢ļxūėĮM║Ž═Č╝ė>65%Ż¼ūŅ╝č95%(10 mmol Īż L -1 CaCl 2 + 0.26 mmol Īż L -1 FeCl 3 ) | Kim et al ., 2011 |
| ┐▌▓▌č┐µ▀ŚUŠ· ( Bacillus subtilis ) |
═Č╝ėĘųļx╝ā╗»║¾Ą─░¹═Ō╠ß╚Ī╬’(”├-Š█╣╚░▒╦ß) | įŁ╩╝ąĪŪ“įÕ( Chlorella protothecoides )Ż¼įÕę║ØŌČ╚1.2 g Īż L -1 | ═Č╝ė┴┐10Ī½30 mg Īż L -1 ; pH 6.5Ī½8.5 | ═Č╝ė┴┐10~20 mg Īż L -1 Ż║ą§─²ą¦╣¹ļS═Č╝ė┴┐į÷╝ėČ°į÷ÅŖŻ¼20 mg Īż L -1 Ģr▀_90%Ż╗└^└mį÷┤¾═Č╝ė┴┐ą§─²ą¦╣¹Ž┬ĮĄŻ╗pH¤o’@ų°ė░Ēæ | Zheng et al ., 2012 |
| č┐µ▀ŚUŠ·( Bacillus sp.) | ═Č╝ė╬ó╔·╬’╝Ü░¹(100▒ČØŌ┐s) | ╬óöMŪ“įÕ( Nannochloropsis sp.)Ż¼įÕę║ØŌČ╚1Ī┴10 7 cells Īż mL -1 |
╬ó╔·╬’╝Ü░¹/įÕ╝Ü░¹Ż║ 1/125Ī½25/1Ż╗pH 6Ī½10Ż╗Č■ārĻ¢ļxūė(Ca 2+ /Mg 2+ )0.125Ī½16 mmol Īż L -1 | ╬ó╔·╬’╝Ü░¹/įÕ╝Ü░¹Ż║<1Ģrą§─²ą¦╣¹ļS═Č╝ė┴┐į÷┤¾Č°į÷ÅŖŻ¼=1ĢrūŅ┤¾▀_73%Ż¼└^└mį÷┤¾═Č╝ė┴┐ą§─²ą¦╣¹Ž┬ĮĄŻ╗pH<9ĢrĮŌą§Ż╗Č■ārĻ¢ļxūėĄ─┤µį┌Š▀ėąĻPµIū„ė├Ż¼ęįCa 2+ Ė³×ķ’@ų° | Powell et al ., 2013 |
| Solibacillus silvestris (┼ÓBė┌▓╗═¼╠╝į┤) | ═Č╝ė│ķ╚Īę║(6000 r Īż min -1 ļxą─║¾Ą─╔ŽŪÕę║) | ╬óöMŪ“įÕ( Nannochloropsis oceanica )Ż¼įÕę║ØŌČ╚╬┤ų¬ | ą§─²ä®┴┐/įÕę║┴┐Ż║3:1Ż╗pH 6.7Ī½10.7Ż╗Ļ¢ļxūė(KClŻ¼CaCl 2 Ż¼FeCl 3 )0.01Ī½0.1 mmol Īż L -1 |
pH<8Ż¼ą§─²ą¦╣¹<20%Ż╗pH>8Ż¼75.4%Ī½88.2%Ż╗Ļ¢ļxūė¤oė░ĒæŻ╗╠╝į┤ĘNŅÉėą’@ų°ė░Ēæ | Wan et al ., 2013 |
▒Ē 3 Ė„ĘN╔·╬’ą§─²ä®į┌╬óįÕĘųļx▓╔╩šųąĄ─æ¬ė├
ĪĪĪĪį┌ą§─²ÖC└Ē╔ŽŻ¼Į^┤¾▓┐Ęų╬─½Iųąų╗┐╝æ]┴╦ņoļŖū„ė├┴”Ż¼░č╔·╬’ą§─²ÜwĮYė┌╬³ĖĮ╝▄ś“ū„ė├Ż¼Š▀¾wėų┐╔╝ÜĘų×ķęįŽ┬ā╔ĘNÖCųŲŻ║ó┘ķLµ£EPSį┌▓╗═¼▓┐╬╗╬³ĖĮČÓéĆĦžōļŖĄ─įÕ╝Ü░¹ą╬│╔╝▄ś“ū„ė├(Bridging); ó┌Č╠µ£EPSį┌Šų▓┐─µ▐DįÕ╝Ü░¹Ą─ļŖžōąįŻ¼Å─Č°ą╬│╔╦∙ų^Ą─ņoļŖ╗źčaą¦æ¬(Patching)(łD 3).▀@ę╗└ĒšōĄ─╗∙ĄA╩Ūīó╔·╬’ą§─²ä®(EPS)─¼šJ×ķĻ¢ļxūėą═Ė▀Š█╬’. Ą½╚ń2.2.1╣Ø╦∙╩÷Ż¼EPSį┌ųąąį╝░ēAąįŚl╝■Ž┬▒Š╔Ē╩ŪĦžōļŖĄ─. ─Ū├┤Ż¼╔·╬’ą§─²ä®╩Ū╚ń║╬īŹ¼FĻ¢ļxūė╗»Ą──ž?ę╗éĆŽÓī”│╔╩ņĄ─└Ēšō×ķČ■ārĻ¢ļxūė╝▄ś“└Ēšō(Divalent Cation Bridging(DCB)Theory)Ż¼┐╔ĮY║ŽłD 4šf├„╚ńŽ┬: EPS▒Š╔ĒŠ▀ėąČÓéĆĦžōļŖĄ─╗Ņąį▓┐╬╗Ż¼▀@╩╣Ųõ─▄ÅŖ┴ę╬³ĖĮŁhŠ│ųąĄ─Č■ārĻ¢ļxūė. ▒╗╬³ĖĮĄ─Č■ārĻ¢ļxūė╦∙Ħš²ļŖ║╔ų╗▒╗EPSųą║═┴╦ę╗░ļŻ¼╦∙ęį─▄┴Ē═Ō╬³ĖĮę╗éĆĦžōļŖĄ─įÕ╝Ü░¹. ė╔┤╦Ż¼ČÓéĆįÕ╝Ü░¹═©▀^Č■ārĻ¢ļxūėĄ─╝▄ś“ū„ė├▀BĮėį┌EPS╔ŽŻ¼ą╬│╔┤¾Ą─ą§¾w. ▀@ę╗└Ēšō─▄║▄║├ĄžĮŌßī×ķ╩▓├┤ČÓārĻ¢ļxūėī”╔·╬’ą§─²Š▀ėą’@ų°Ą─ÅŖ╗»ū„ė├╔§ų┴╩Ūą§─²ą╬│╔Ą─▒žę¬Śl╝■. ▀@ę╗└Ēšōę▓─▄ĮŌßīą§─²ą¦╣¹ļSpH╔²Ė▀Č°į÷ÅŖĄ─¼FŽ¾: pH╔²Ė▀Ż¼EPSļŖžōąįį÷ÅŖŻ¼╬³ĖĮČ■ārĻ¢ļxūėĄ──▄┴”į÷ÅŖŻ¼╦∙ęį╝▄ś“ū„ė├į÷ÅŖ. Ą½╚ń╔Ž╦∙╩÷Ż¼į┌▓┐Ęų蹊┐ųąĻ¢ļxūėĄ─┤µį┌ī”ą§─²ą¦╣¹Ė∙▒ŠŠ═ø]ėąė░Ēæ(Wan et al., 2013). ▀@Š═¤oĘ©ė├DCB└ĒšōĮŌßī┴╦. DCB└ĒšōūŅĖ∙▒ŠĄ─╚▒Ž▌į┌ė┌Ż¼└Ēšō╔Žæ¬įō╩Ū╦∙ėąÄ¦žōļŖĄ─EPSČ╝─▄═©▀^Č■ārĻ¢ļxūėĄ─╝▄ś“ū„ė├ą╬│╔ą§─². Ą½▀@’@╚╗╩Ū┼c╩┬īŹ▓╗Ę¹Ą─. ▀@ę╗å¢Ņ}īóį┌4.3╣Ø▓┐ĘųĄ├ĄĮ▀Mę╗▓Įėæšō.
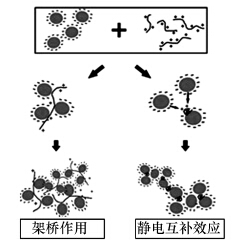
ĪĪĪĪłD 3 ╔·╬’ą§─²ä®Ą─ą§─²ÖC└Ē(Salim et al., 2011)
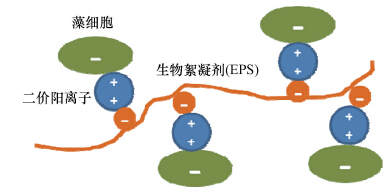
ĪĪĪĪłD 4 Č■ārĻ¢ļxūė╝▄ś“įŁ└Ē
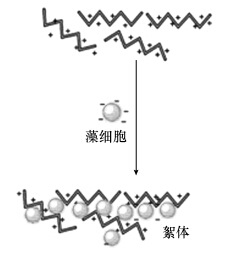
ĪĪĪĪ4 ╬óįÕūį░ląįą§─² 4.1 Ė┼╩÷
ĪĪĪĪ╬óįÕėąĢr─▄į┌ø]ėą╠Ē╝ė╚╬║╬ą§─²ä®Ą─ŪķørŽ┬░l╔·ą§─²Ż¼▀@ę╗¼FŽ¾▒╗ĘQ×ķūį░ląįą§─²(Auto-flocculation)╩ūŽ╚├Ķ╩÷┴╦▀@ę╗¼FŽ¾. ╦¹éā░l¼FŻ║įÕŅÉ╠┴ųąĄ─╬óįÕį┌£žČ╚▌^Ė▀Ūę╣ŌŠĆ│õūŃĄ─Ģr║“─▄ūį╚╗Ąžą╬│╔ą§¾w.┤╦║¾Ż¼║▄ČÓīWš▀Č╝ūCīŹ┴╦ŅÉ╦Ų¼FŽ¾Ą─┤µį┌▓óķ_š╣┴╦ŽÓĻP蹊┐. ─┐Ū░ą╬│╔Ą─╗∙▒Š╣▓ūR×ķŻ¼╬óįÕūį░ląįą§─²╩Ūė╔ā╔ĘN▓╗═¼ÖC└Ēę²░lĄ─Ż║
ĪĪĪĪ(1)į┌Ė▀pHŽ┬Ż¼Ō}ĪóµVĄ╚ļxūėą╬│╔Ħš²ļŖĄ─│┴ĄĒ╬’Ż¼ŲĄĮļŖąįųą║═ū„ė├Å─Č°ę²░lą§─². ╬─½IųąĄ─ūį░ląįą§─²ę╗░Ń╝┤ųĖ┤╦ŅÉ. Ė▀pH┐╔ęį╩Ūė╔╬óįÕ╣Ō║Žū„ė├Ž¹║─╦«ųą¤oÖC╠╝(Inorganic CarbonŻ¼IC)ūį╚╗ą╬│╔Ż¼ę▓┐╔═©▀^╚╦╣ż╠Ē╝ėēAąį╬’┘|(╩»╗ęŻ¼Üõč§╗»ŌcĄ╚)Č°ą╬│╔. ć└Ė±üĒšfŻ¼ų╗ėąŪ░š▀▓┼Ę¹║Žūį░ląįą§─²Ą─Č©┴x. Ą½┐╝æ]ĄĮā╔š▀Ą─īŹ┘|Č╝╩Ūą╬│╔Ħš²ļŖĄ─│┴ĄĒ╬’Ż¼▒Š╬─į┌┤╦īóā╔š▀ę╗▓ó╝{╚ļĖ▀pHšTī¦Ą─ūį░ląįą§─²ĘČ«Ā.
ĪĪĪĪ(2)▓┐ĘųįÕĘNį┌Ųõ╔·└Ē╗Ņäėųą─▄«a╔·┤¾┴┐Š▀ėąą§─²ū„ė├Ą─░¹═ŌŠ█║Ž╬’Ż¼ŲĄĮ╔·╬’ą§─²ä®Ą─ū„ė├(ģóęŖ3.3╣Ø▓┐Ęų)Å─Č°ę²░lą§─². Ė▀ą¦įÕŅÉ╠┴ųą│ŻęŖĄ─╝»ąŪįÕī┘(Actinastrum)Īó╬ó├óįÕī┘(Micractinium)Īó¢┼įÕī┘(Scenedesmus)Īó┐šąŪįÕī┘(Coelastrum)Īó▒PąŪįÕī┘(Pediastrum)╝░─zŠWįÕī┘(Dictyosphaerium)Ą╚│Ż═©▀^įōÖC└Ēą╬│╔┤¾Ą─╚║┬õĮYśŗ(50~200 ”╠m)Č°Ą├ęįūį╚╗│┴ĮĄ. ╬─½Iųą│ŻīóŲõÜw×ķ╔·╬’ą§─²ę╗ŅÉ(ģóęŖ3.3╣Ø▓┐Ęų). ę“×ķįōŪķørŽ┬Ą─╔·╬’ą§─²ä®×ķįÕ╝Ü░¹ūį╔Ē╦∙«a╔·Ż¼▒Š╬─į┌┤╦ę▓īóŲõ╝{╚ļūį░ląįą§─²▓óČ©┴x×ķEPSę²ŲĄ─ūį░ląįą§─².
ĪĪĪĪ4.2 Ė▀pHšTī¦Ą─ūį░ląįą§─²
ĪĪĪĪ╚ń╔Ž╦∙╩÷Ż¼Ė▀pHšTī¦Ą─ūį░ląįą§─²ŲõīŹ┘|×ķ╦∙╔·│╔Ą─Ħš²ļŖ│┴ĄĒ╬’Ą─ļŖąįųą║═ū„ė├. ę“┤╦Ż¼šTī¦┤╦ŅÉūį░ląįą§─²Ą─ĻPµIŠ═╩Ū├„┤_į┌╬óįÕš²│Ż┼ÓBŚl╝■Ž┬╦∙─▄ą╬│╔Ą─│┴ĄĒ╬’ĘNŅÉ║═ąį┘|. ╬█╦«ųąę╗░Ń║¼ėą┤¾┴┐Ą─Ō}ĪóµVĪó╠╝╦ßĖ∙║═┴ū╦ßĖ∙Ą╚ļxūėŻ¼į┌ēAąįŚl╝■Ž┬▒╚▌^╚▌ęūą╬│╔Ą─│┴ĄĒ╬’ų„ę¬░³└©Ż║┴ū╦ßŌ}Ż¼Üõč§╗»µV║═╠╝╦ßŌ}. ┤¾┴┐蹊┐ūCīŹŻ¼╠╝╦ßŌ}▒Š╔ĒĦžōļŖŻ¼ūŅČÓų╗─▄═©▀^ŠW▓Č║═ŠĒÆ▀ū„ė├īŹ¼FĘŪ│ŻėąŽ▐Ą─ą§─²ą¦╣¹. ┴ū╦ßŌ}║═Üõč§╗»µVĦš²ļŖŻ¼└Ēšō╔ŽČ╝┐╔šTī¦ūį░ląįą§─². Ą½ā╔š▀ą╬│╔Ą─Š▀¾wŚl╝■▓Ņäe▌^┤¾Ż¼ęįų┬╬─½IųąĄ─ĮYšō│Ż│Ż╚▌ęūę²Ųš`ĮŌ. ×ķ┤╦Ż¼ęįŽ┬ėĶęįįö╩÷.
ĪĪĪĪSukenik║═Shelef(1984)╩ū┤╬Č©┴┐ĪóŽĄĮyąįĄžčąŠ┐┴╦Ė▀pHŽ┬Ą─╬óįÕūįą§─²¼FŽ¾Ż¼ĮYšōąįšJ×ķŻ¼┴ū╦ßŌ}╩ŪšTī¦ūįą§─²Ą─ĻPµI│┴ĄĒ╬’.╦¹éāĄ─įć“×░³└©ā╔▓┐ĘųŻ║ó┘æ¶═Ō┼ÓBŽĄĮyĄ─ūįą§─²įć“×: ▓╔ė├ķgą¬┼ÓBŻ¼ų▄Ų┌×ķ8 d; Ū░7 d▀B└m═©╚ļCO2╩╣pHŠS│ųį┌7.0; Ą┌8╠ņ═Żų╣CO2╣®æ¬║═öć░ĶŻ¼▒O£yą§─²ą¦╣¹║═ŽÓĻP╦«┘|ģóöĄ(pHŻ¼TSSŻ¼PO43--PŻ¼Ca2+Ż¼Mg2+Ż¼ēAČ╚)Ą─ūā╗»Ūķør. ó┌╩ęā╚ą§─²įć“×(Jar Test): ╚Ī╠Äė┌ī”öĄį÷ķLŲ┌Ą─Č■ą╬¢┼įÕ║═ąĪŪ“įÕų├ė┌║═æ¶═Ō┼ÓBŽĄĮyŽÓ═¼Ą─┼ÓB╗∙Ż¼š{╣ØpH×ķ2.5Ī½10.5▀Mąąą§─²įć“×(80 r Īż min-1 1 minŻ¼30 r Īż min-1 15minŻ¼│┴ĄĒ15 min)Ż¼▒O£yą§─²ą¦╣¹║═ŽÓĻP╦«┘|ģóöĄūā╗». æ¶═Ō┼ÓBįć“×ĮY╣¹▒Ē├„Ż¼ŽĄĮy═Żų╣CO2╣®æ¬║═öć░Ķ24 h║¾Ż¼¾wŽĄpHčĖ╦┘╔²ų┴8.9Ż¼PO43--PĪóCa2+═¼▓ĮĮĄĄ═Ż¼Ė▀ą¦ą§─²(96%)ą╬│╔Ż¼Č°Mg2+ŠS│ų▓╗ūā(▒Ē 4). ė╔┤╦│§▓ĮūC├„Ż¼ūį░ląįą§─²╩Ūė╔Ė▀pHŽ┬Ą─┴ū╦ßŌ}│┴ĄĒšTī¦Ą─. ╩ęā╚ą§─²įć“×ĮY╣¹’@╩ŠŻ║pH 5.0Ī½7.5Ģr¤oą§─²░l╔·. pH>8.5║¾ą╬│╔┴╦Ė▀ą¦ą§─²(Ī½98%)Ż¼┼cæ¶═Ōįć“×ĮY╣¹ę╗ų┬. ▀Mę╗▓Įįć“×’@╩ŠŻ¼į┌PO43--P×ķ6.2 mg Īż L-1Ą─ŪķørŽ┬Ż¼Ca2+(2.0 mmol Īż L-1)į┌pHĪ▌8.5ĢrŠ═─▄╔·│╔│┴ĄĒ▓óę²░lą§─²;Č°Mg2+(2.0 mmol Īż L-1)ę¬į┌pHĪ▌10.5║¾▓┼─▄╔·│╔│┴ĄĒ▀MČ°ę²░lą§─². ╚ń╣¹ø]ėąPO43--P┤µį┌Ż¼ätCa2+į┌š¹éĆēAąįĘČć·ā╚Č╝▓╗─▄ę²Ųą§─².
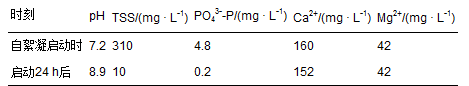
ĪĪĪĪ▒Ē 4 ūįą§─²Ū░║¾Ą─╦«┘|ģóöĄūā╗»
ĪĪĪĪ▀@ą®īŹ“×ĮY╣¹│õĘųūC├„Ż¼į┌╬óįÕ╔·ķLĄ─š²│ŻpHĘČć·ā╚(8Ī½10)Ż¼┴ū╦ßŌ}╩ŪšTī¦ūįą§─²Ą─ĻPµI│┴ĄĒ╬’. Ė³Š▀╣ż│╠ęŌ┴xĄ─╩ŪŻ¼Sukenik║═Shelef(1984)░l¼Fūįą§─²Ą─┼RĮńpHųĄ(ą§─²ą¦╣¹▀_50%Ą─pH)īóļSPO43-(▒Ē 5)║═Ca2+(╬┤’@╩Š)ØŌČ╚Ą─╔Ž╔²Č°Ž┬ĮĄ; į┌║¼Ė▀ØŌČ╚Ca2+║═PO43- Ą─┼ÓBę║ųąŻ¼╬óįÕūįą§─²╔§ų┴┐╔ęįį┌ųąąįŚl╝■Ž┬Š═░l╔·. ▀@šf├„«öęį║¼┴ū▌^Ė▀Ą─╩ąš■╬█╦«×ķĄū╬’ĢrŻ¼┐╔ŽÓī”▌^╚▌ęūĄžį┌╬óįÕ╔·ķLĄ─š²│ŻpHųĄĘČć·ā╚Ż¼═©▀^╔·│╔┴ū╦ßŌ}│┴ĄĒ═¼▓ĮīŹ¼Fūį░ląįą§─²║═Ė▀ą¦│²┴ū.
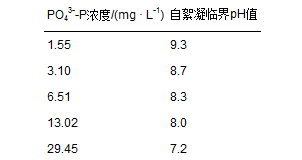
ĪĪĪĪ▒Ē 5 ąĪŪ“įÕį┌▓╗═¼┴ūØŌČ╚Ž┬Ą─ūįą§─²┼RĮńpHųĄ
ĪĪĪĪ╚╗Č°Ż¼│²╔┘öĄīWš▀└^└mūCīŹ┴ū╦ßŌ}į┌šTī¦╬óįÕūįą§─²ųąĄ─║╦ą─ū„ė├═ŌŻ¼┤¾▓┐Ęų蹊┐Č╝’@╩ŠÜõč§╗»µV▓┼╩ŪšTī¦ūįą§─²Ą─ĻPµI│┴ĄĒ╬’Ż¼┤·▒Ēąį蹊┐╚ńŽ┬Ż║
ĪĪĪĪ(1)VandammeĄ╚(2012a)į┌蹊┐ąĪŪ“įÕūįą§─²Ģr░l¼FŻ¼į┌pHĪ▄10.5ĢrŠ∙¤oūįą§─²░l╔·; «öpHš{ų┴11ĢrŻ¼īŹ¼F┴╦75%Ą─ą§─²ą¦╣¹; pHĪ▌11.5║¾ą§─²ą¦╣¹’@ų°Ż¼▀_95%ęį╔Ž(▒Ē 6). ┤╦Ģr╚¶╝ė╚ļEDTA(ūŅĮKØŌČ╚0.5M)č┌▒╬Ca2+ĪóMg2+Ą╚Č■ārļxūėŻ¼ätą§─²ą¦╣¹╝▒äĪŽ┬ĮĄų┴<20%. ▀@│§▓ĮūC├„┴╦Ō}/µV│┴ĄĒį┌šTī¦ūįą§─²ųąĄ─ĻPµIū„ė├. ▀Mę╗▓Į蹊┐▒Ē├„Ż¼╚¶╚▄ę║ųąų╗┤µį┌Ca2+(1~100 mg Īż L-1)Ż¼į┌Ė▀pHŽ┬(10.5~12)┤_īŹę▓─▄ą╬│╔│┴ĄĒ╬’Ż¼Ą½╩╝ĮK▓╗─▄ą╬│╔ėąą¦ą§─²(<20%). Č°«ö╚▄ę║ųą┤µį┌Mg2+(1.8 mg Īż L-1)ĢrŻ¼į┌pH=10.5Š═┐╔īŹ¼F25%Ą─ą§─²ą¦╣¹; į┌pH=12Ģrą§─²ą¦╣¹’@ų°į÷ų┴85%. «öMg2+ØŌČ╚Ī▌3.6 mg Īż L-1║¾Ż¼pH=10.5Š═┐╔▀_90%ęį╔ŽĄ─ą§─²ą¦╣¹(ūóŻ║įōīŹ“׌l╝■(įÕ╝Ü░¹ØŌČ╚Ą╚)┼c▒Ē 6▓╗═¼Ż¼╣╩┼RĮńpH║═ą§─²ą¦╣¹ėą╦∙▓Ņ«É).
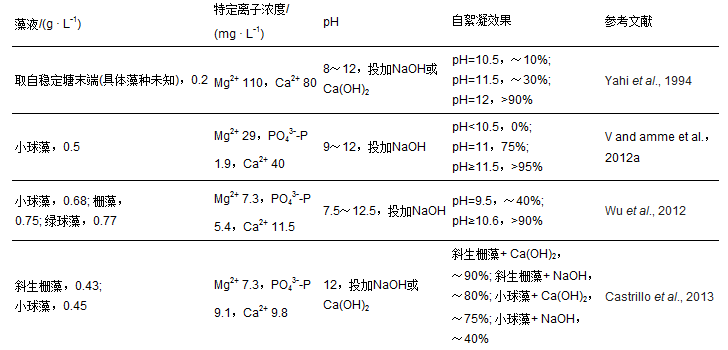
ĪĪ▒Ē 6 ęįÜõč§╗»µV│┴ĄĒšTī¦Ą─ūį░ląįą§─²
ĪĪĪĪ(2)SirinĄ╚(2012)į┌蹊┐╚²ĮŪ║ųųĖįÕĄ─ūįą§─²Ģr░l¼FŻ¼į┌pH=10.5Ģrą§─²║¾╚▄ę║ųąĄ─Mg2+Ž┬ĮĄ┴╦╝s16%Ż¼Č°Ca2+╗∙▒Š▓╗ūā; «öpHį÷ų┴11ĢrŻ¼╚▄ę║ųą╗∙▒Šęčø]ėąMg2+Üł┴¶Ż¼Č°Ca2+āHĮĄĄ═┴╦14%(Sirin et al., 2012). ▀@šf├„į┌pH=10.5Ī½11Ģr╔·│╔Ą─│┴ĄĒ╬’ų„ę¬×ķµV│┴ĄĒ╬’.
ĪĪĪĪ(3)Smith║═Davis(2012)░l¼FŻ¼Mg2+ĪóCa2+║═CO32-ļxūėųąŻ¼ų╗ėąMg2+(9.6 mmol Īż L-1)┤µį┌Ģr▓┼─▄į┌Ė▀pHŽ┬(>10)īŹ¼FĖ▀ą¦ą§─²;Č°Mg2+╚▒Ę”Ģr(Ca2+ĪóCO32-Š∙×ķ9.6 mmol Īż L-1)Ż¼╝┤╩╣Ca2+│┴ĄĒ┴╦75%ę▓▓╗─▄ą╬│╔ėąą¦ą§─²(Smith and DavisŻ¼ 2012).
ĪĪĪĪęį╔ŽīŹ“×ĮY╣¹╦Ų║§Š∙ęčūC├„Ż¼Ė▀pHŽ┬ų╗ėąMg│┴ĄĒ╬’▓┼─▄ėąą¦šTī¦ūįą§─². ╚╗Č°Ż¼ūą╝Ü┐╝▓ņ▀@ą®čąŠ┐ųąĄ─╠žČ©ļxūėØŌČ╚(▒Ē 6)Š═┐╔░l¼FŻ¼ŲõCa2+ĪóPO43-ļxūėųąų┴╔┘ėąę╗ĒŚ╗“ā╔ĒŚĄ─ØŌČ╚Š∙▌^Ą═(┼c▒Ē 5ī”▒╚)Ż¼ę“┤╦Ż¼į┌pH=8Ī½10Ą─ĘČć·ā╚▓╗─▄ą╬│╔┤¾┴┐┴ū╦ßŌ}│┴ĄĒ; ų╗─▄į┌Ė³Ė▀pHŽ┬(>10.5)╔·│╔Mg(OH)2│┴ĄĒĢr▓┼─▄šTī¦ūįą§─². ŠC║Žęį╔Ž╦∙ėąĮY╣¹Ż¼┐╔ęįĄ├│÷╚ńŽ┬ĮYšō: ┴ū╦ßŌ}║═Üõč§╗»µV│┴ĄĒČ╝┐╔ėąą¦šTī¦│÷ūį░ląįą§─². PO43--PĪóCa2+ØŌČ╚Š∙▌^Ė▀ĢrŻ¼┴ū╦ßŌ}│┴ĄĒį┌ŽÓī”▌^╚§Ą─ēAąįŚl╝■Ž┬(pH 8~10)Š═┐╔╔·│╔▓óšTī¦│÷’@ų°Ą─ūįą§─²;PO43--P/Ca2+ØŌČ╚▌^Ą═ĢrŻ¼ätąĶ▀Mę╗▓Į╠ß╔²pHų┴10.5ęį╔ŽŻ¼«a╔·Üõč§╗»µV│┴ĄĒ║¾▓┼─▄šTī¦│÷ūįą§─².
ĪĪĪĪ4.3 ░¹═ŌŠ█║Ž╬’(EPS)ę²ŲĄ─ūį░ląįą§─²
ĪĪĪĪ20ČÓ─ĻęįŪ░Ż¼╝«EPSą╬│╔ūį░ląįą§─²Ą─įÕĘN▒Ńį┌╬óįÕĘųļx▓╔╩šųąĄ├ĄĮųžęĢ.Borowitzka║═BorowitzkaĘųļx│÷┴╦╦{įÕķTĄ─ę╗ųĻ─zŪ╩įÕ. ╦³─▄Ęų├┌│÷┤¾┴┐Š▀ėąą§─²ū„ė├Ą─EPSŻ¼Ųõų„ę¬│╔Ęų░³└©ČÓŠ█╠ŪĪóų¼ĘŠ╦ß║═Ą░░ū┘|.┤╦║¾Ż¼ūįą§─²įÕĘN╚ń§U╩ŽŽ»╬óįÕ╝░ĮzĀŅįÕĄ╚į┌╬█╦«╠Ä└ĒųąČ╝Ą├ĄĮ┴╦æ¬ė├. Ą½Ģrų┴Į±╚šŻ¼ī”┤╦ŅÉūįą§─²Ą─蹊┐┐é¾wüĒšf▀Ć╩«ĘųėąŽ▐.
ĪĪĪĪį┌īŹļHæ¬ė├╔ŽŻ¼ę╗░Ń╦╝┬Ę×ķīóūįą§─²įÕĘN═Č╚ļĘŪūįą§─²įÕĘN┼ÓBŽĄĮyęįīŹ¼Fą§─²Ęųļx. SalimĄ╚(2011)蹊┐┴╦ĄŁ╦«ūįą§─²įÕĘNńĀą╬└wŠSįÕą▒╔·¢┼įÕī”ąĪŪ“įÕą§─²ū„ė├╝░║Żč¾ūįą§─²įÕĘN╦─▒▐Ų¼įÕī”Ė╗ė═ą┬ŠGįÕĄ─ą§─²ū„ė├.īŹ“×ĮY╣¹’@╩ŠŻ¼╦─▒▐Ų¼įÕĄ─ą§─²ą¦╣¹ūŅ╝čŻ¼┐╔▀_70%ū¾ėę;ą▒╔·¢┼įÕ┤╬ų«Ż¼ą§─²ą¦╣¹┐╔▀_30%ū¾ėę;ńĀą╬└wŠSįÕą§─²ą¦╣¹ūŅ▓ŅŻ¼╝s20%. GuoĄ╚┐╝▓ņ┴╦ūįą§─²ą▒╔·¢┼įÕįÕųĻS. obliquus AS-6-1ī”ĘŪą§─²ĄŁ╦«įÕųĻS. obliquus FSP-3ĪóC. vulgaris CNW-11║═║Żč¾╬óįÕN. oceanica DUT01Ą─ą§─²ū„ė├. S. obliquus AS-6-1ī”ĄŁ╦«įÕųĻŠ∙╚ĪĄ├┴╦80%ęį╔ŽĄ─┴╝║├ą§─²ą¦╣¹;Č°ī”║Żč¾╬óįÕĄ─ą§─²ą¦╣¹ätŽÓī”▌^▓ŅŻ¼Ą═ė┌60%.ęį╔ŽīŹ“×ĮY╣¹▒Ē├„Ż¼ūįą§─²įÕųĻĄ─ą§─²ą¦╣¹Ģ■ę“─┐ś╦įÕĘNČ°«ÉŻ¼▀@į┌īŹļHæ¬ė├ųą┤µį┌ų°║▄┤¾Ą─ŠųŽ▐ąį.
ĪĪĪĪ─┐Ū░ī”ė┌╬óįÕEPSšTī¦ūįą§─²Ą─ÖC└ĒĄ─蹊┐Ė³×ķėąŽ▐Ż¼ę╗░Ńų╗╩Ū╗\ĮyĄžšJ×ķ┼c╔·╬’ą§─²ä®Ą─ÖC└Ēę╗ų┬(ģóęŖ3.3╣Ø▓┐Ęų). į┌ŽÓ╗źū„ė├┴”╔ŽŻ¼Į^┤¾▓┐Ęų蹊┐ų╗┐╝æ]┴╦╗∙ė┌DCBįŁ└ĒĄ─ņoļŖū„ė├┴”Ż¼ų╗ėąūŅĮ³śO╔┘öĄčąŠ┐┐╝æ]┴╦Lewis╦ß-ēA╦«║Žū„ė├┴”. ╚ń3.3╣Ø╦∙╩÷Ż¼å╬╝āĄžė├DCBįŁ└Ē(łD 4)ĮŌßīįÕ╝Ü░¹EPSĄ─ą§─²ÖC└Ēīó┤µį┌║▄┤¾Ą─╚▒Ž▌. ę“×ķ░┤┤╦└Ēšō╦∙ėą«a╔·EPSĄ─╬óįÕĘNī┘Č╝æ¬įō─▄═©▀^DCBįŁ└Ē░l╔·ūįą§─²Ż¼Č°īŹļH╔Žų╗ėą─│ą®╠žČ©įÕĘN«a╔·Ą─EPS▓┼ėąą§─²ū„ė├. ę╗éĆ┐╔─▄Ą─ĮŌßī×ķŻ║ĘŪą§─²įÕĘN«a╔·Ą─EPSöĄ┴┐▌^╔┘Ż¼╝▄ś“─▄┴”ėąŽ▐;Č°ūįą§─²įÕĘN─▄«a╔·┤¾┴┐EPSŻ¼╦∙ęįą§─²ą¦╣¹’@ų°. ┴Ēę╗éĆ┐╔─▄Ą─ĮŌßī×ķŻ¼ĘŪą§─²įÕĘN║═ūįą§─²įÕĘN╦∙«a╔·Ą─EPSį┌ĮM│╔║═ąį┘|╔Žėą╦∙▓╗═¼. └²╚ńŻ¼GuoĄ╚(2013)░l¼FŻ¼į┌EPSĖ„ĮMĘųųąų╗ėąČÓŠ█╠Ū×ķą§─²Ą─╗Ņąį│╔Ęų.ę“┤╦Ż¼╚ń╣¹▓╗═¼įÕĘN«a╔·Ą─EPSųąČÓŠ█╠ŪĄ─║¼┴┐▓╗═¼Ż¼─ŪŲõą§─²ą¦╣¹Š═┐╔─▄ėą’@ų°▓Ņ«É. Ą½GuoĄ╚▀Mę╗▓Į░l¼FŻ║¤ošō╩Ū═Č╝ėCa2+Ż¼▀Ć╩Ūė├EDTAč┌▒╬Ca2+Ż¼ī”ą§─²ą¦╣¹Č╝ø]ėąė░ĒæŻ¼▀@Š═Å─Ė∙▒Š╔Ž┼cDCB└ĒšōŽÓŃŻ┴╦.
ĪĪĪĪOzkan║═BerberogluätÅ─įÕ╝Ü░¹Ą─ėH/╩Ķ╦«ąį▒Ē├µ╠žąį│÷░lŻ¼į┌ņoļŖū„ė├┴”Ą─╗∙ĄA╔Žį÷╝ė┴╦ī”įÕ╝Ü░¹ķgLewis╦ß-ēA╦«║Žū„ė├┴”Ą─┐╝▓ņ. ╦¹éāĄ─蹊┐ĮY╣¹▒Ē├„: Lewis╦ß-ēA╦«║Žū„ė├┴”į┌XDLVOĄ─3ĘN╗∙▒Šū„ė├┴”ųąūŅÅŖŻ¼į┌╬óįÕĄ─ūįą§─²ųąŠ▀ėąĻPµIū„ė├. «ö╬óįÕæęĖĪę║ųą┤µį┌▀m┴┐╩Ķ╦«ąį▌^ÅŖĄ─╬óįÕĘNī┘(╚ńŻ¼▓╝└╩ŲŽ╠čįÕ)ĢrŻ¼╝┤╩╣╩ŪļŖžōąį▌^┤¾Ą─ėH╦«-╩Ķ╦«╗ņ║ŽįÕę║ę▓─▄ą╬│╔ą§─²; ╚ń╣¹ų╗┐╝æ]ĘČĄ┬╚A┴”║═ņoļŖ┴”Ż¼▀@ę╗¼FŽ¾īó¤oĘ©Ą├ĄĮĮŌßī(GTOT(d)= GLW(d)+ GEL(d)> 0Ż¼└Ēšō╔Ž▓╗░l╔·ą§─²); Č°┐╝æ]Lewis╦ß-ēA╦«║Žū„ė├┴”║¾Ż¼└ĒšōŅA£y┼cīŹļHė^▓ņĄ├ĄĮ┴╦║▄║├Ą─╬Ū║Ž(GTOT(d)= GLW(d)+ GEL(d)+ GAB(d)< 0Ż¼└ĒšōŅA£y×ķ░l╔·ą§─²). ▀@ą®čąŠ┐│╔╣¹×ķ└ĒĮŌEPSšTī¦ūįą§─²Ą─ÖC└Ē║═┤┘▀Mūįą§─²ą¦╣¹╠ß╣®┴╦ĘŪ│ŻėąŪ░Š░Ą─╦╝┬Ę. ę“┤╦Ż¼║¾└m¤ošōī”╔·╬’ą§─²▀Ć╩ŪEPSšTī¦Ą─ūįą§─²Ż¼Č╝æ¬į┌═Ļš¹Ą─XDLVO└Ēšō┐“╝▄ā╚┐╝▓ņĘČĄ┬╚A┴”ĪóņoļŖ┴”║═Lewis╦ß-ēA╦«║Žū„ė├┴”Ą─ŠC║Žū„ė├. ī”╗Ņąį╬█─ÓEPSĄ─蹊┐░l¼F: EPSųąĄ─Ą░░ū┘|╩Ūą╬│╔╩Ķ╦«ąįĄ─ų„ę“Ż¼Č°╠╝╦«╗»║Ž╬’╩Ūą╬│╔ėH╦«ąįĄ─ų„ę“; EPSĄ─öĄ┴┐║═ĮM│╔╩▄╔·ķLļAČ╬ĪóĄū╬’╦«ŲĮĄ╚ę“╦žė░ĒæŻ¼ī”╬ó╔·╬’ą§─²ėąĻPµIė░Ēæ.Č°ī”įÕ╝Ü░¹EPSĖ„ĮMĘųĄ─«a╔·Īóūā╗»ęÄ┬╔╝░Ųõī”▒Ē├µ╠žąįė░ĒæĄ─ŽĄĮy蹊┐Äū║§▀Ć╩Ū┐š░ū. ▀@ĘĮ├µĄ─蹊┐¤oę╔īó×ķ└ĒĮŌ║═š{┐ž╬óįÕūįą§─²╠ß╣®ĘŪ│ŻėąārųĄĄ─ą┼Žó.
ĪĪĪĪ4.4 ė░Ēæę“╦ž
ĪĪĪĪūį░ląįą§─²Ą─░l╔·ÖC└ĒøQČ©┴╦Ųõė░Ēæę“╦ž. ī”ė┌Ė▀pHšTī¦Ą─ūį░ląįą§─²Ż¼ŲõĖ∙▒Šę“╦ž×ķ╬óįÕ╣Ō║Žū„ė├╠ß╔²pH╦∙─▄ĄĮ▀_Ą─│╠Č╚║═╦∙─▄ą╬│╔Ħš²ļŖ│┴ĄĒ╬’Ą─╠žČ©ļxūėØŌČ╚. ī”ė┌EPSę²ŲĄ─ūį░ląįą§─²Ż¼Ųõė░Ēæę“╦žätĖ³╝ėÅ═ļsŻ¼└Ēšō╔Ž░³└©╦∙ėąė░ĒæEPS«a╔·║═ĮM│╔Ą─ę“╦ž.
ĪĪĪĪ4.4.1 ╣Ōšš
ĪĪĪĪ╣Ōšš╩Ū╬óįÕ╔·ķLĘ▒ų│Ą─╗∙▒Šę¬╦žŻ¼ī”Ė▀pH║═EPSšTī¦Ą─ūį░ląįą§─²Č╝Š▀ėąųžę¬ė░Ēæ. ╩ūŽ╚Ż¼╣Ōššų▒ĮėøQČ©┴╦╬óįÕ╣Ō║Žū„ė├Ą─│╠Č╚. ╣ŌššįĮÅŖŻ¼╣Ō║Žū„ė├įĮ│õĘųŻ¼╦«ųą¤oÖC╠╝Ž¹║─įĮÅžĄūŻ¼pH╔Ž╔²įĮĖ▀Ż¼įĮėą└¹ė┌Ė▀pHšTī¦Ą─ūįą§─²░l╔·. Ųõ┤╬Ż¼╣Ōššę▓╩Ūė░ĒæEPS«a╔·Ą─ĻPµIę“ūė. MorenoĄ╚░l¼F╣ŌššÅŖČ╚ė╔345 ”╠mol Īż m-2 Īż s-1į÷╝ėĄĮ460 ”╠mol Īż m-2 Īż s-1║¾Ż¼¶~ą╚įÕĄ─EPS║¼┴┐į÷╝ė┴╦4▒Č. Rebolloso-FuentesĄ╚ę▓░l¼FŻ¼į┌▌^Ė▀Ą─═Ō▓┐╣ŌššŚl╝■Ž┬Ż¼ūŽŪ“įÕEPSĄ─║¼┴┐’@ų°į÷╝ė. ę“┤╦Ż¼│õĘųĄ─╣Ōšš╩ŪšTī¦ūį░ląįą§─²Ą─ėą└¹ę“╦ž.
ĪĪĪĪ4.4.2 ╠žČ©ļxūė
ĪĪĪĪCa2+ĪóMg2+║═PO43-Ą╚╠žČ©ļxūėĄ─ØŌČ╚øQČ©┴╦│┴ĄĒ╬’Ą─ĘNŅÉ║═«a╔·Ą─┼RĮńpHųĄŻ¼ī”Ė▀pHšTī¦Ą─ūį░ląįą§─²Š▀ėąøQČ©ąįė░Ēæ. Å─░l╔·ÖC└ĒüĒ┐┤Ż║PO43--P/Ca2+ļxūėČ╝┤¾┴┐┤µį┌ĢrŻ¼į┌▌^╚§Ą─ēAąįŚl╝■Ž┬(pH 8Ī½10)┴ū╦ßŌ}│┴ĄĒŠ═┐╔┤¾┴┐╔·│╔▓ó│╔×ķūįą§─²ų„ī¦ę“╦ž;PO43--P/Ca2+Ą─Ųõųąų«ę╗ØŌČ╚▌^Ą═ĢrŻ¼ätąĶ▀Mę╗▓Į╠ß╔²pHų┴10.5ęį╔ŽŻ¼«a╔·Üõč§╗»µV│┴ĄĒ║¾▓┼─▄šTī¦│÷ūįą§─²(ģóęŖ4.2╣Ø▓┐Ęų). Å─ą§─²ą¦╣¹üĒ┐┤Ż║ęį╔ŽļxūėØŌČ╚įĮĖ▀Ż¼╔·│╔Ħš²ļŖĄ─│┴ĄĒ╬’įĮČÓŻ¼ļŖąįųą║═─▄┴”įĮÅŖŻ¼ą§─²Š═įĮ│õĘų. ę“┤╦Ż¼īó╔Ž╩÷╠žČ©ļxūėŠS│ųį┌▌^Ė▀╦«ŲĮī”īŹ¼FĖ▀pHšTī¦Ą─ūį░ląįą§─²ų┴ĻPųžę¬.
ĪĪĪĪ4.4.3 £žČ╚║═╔·ķLļAČ╬
ĪĪĪĪ£žČ╚ī”╬óįÕEPSą╬│╔Š▀ėąųžę¬ė░Ēæ.Ė▀£ž┤╠╝żEPSĄ─ą╬│╔Ż¼Č°Ą═£žŽ┬ė╔ė┌╝Ü░¹ą┬ĻÉ┤·ųxĮĄĄ═EPSĄ─ą╬│╔╩▄ĄĮęųųŲ. Ą½EPS«a╔·Ą─ūŅ╝č£žČ╚ę“įÕĘN▓╗═¼Č°«ÉŻ¼╚ńŻ║▓╝└╩ŲŽ╠čįÕ(Botryococcus braunii)į┌£žČ╚Ą═ė┌23 ĪµÄū║§▓╗Ęų├┌EPSŻ¼ŲõūŅ╝č£žČ╚×ķ30Ī½33 ĪµČ°¶~ą╚įÕ(Anabaena sp.)į┌30Ī½35 ĪµĘČć·ā╚EPS«a┴┐Č╝║▄╔┘Ż¼ų╗ėąį┌40 Īµęį╔ŽEPS▓┼┤¾┴┐«a╔·.ĪĪĪĪ╬óįÕ╦∙╠Ä╔·ķLļAČ╬ī”įÕ╝Ü░¹├▄Č╚Īó▒Ē├µąį┘|║═EPSĄ─«a┴┐╝░│╔ĘųĄ╚Č╝ėą’@ų°ė░Ēæ.Lavoie║═de la No©╣e░l¼FŻ¼└Ž²g╗»Ą─(Aging)Ą─įÕ╝Ü░¹├▄Č╚į÷┤¾Ż¼ęūė┌│┴ĄĒ.ZhangĄ╚░l¼FŻ¼ąĪŪ“įÕÅ─ī”öĄį÷ķLŲ┌▀M╚ļĘĆČ©Ų┌║¾Ż¼Ųõ▒Ē├µļŖžōąį£p╚§Ż¼ęūė┌░l╔·ą§─²Lavoie║═de la No©╣eZhangĄ╚║═SalimĄ╚Č╝░l¼F╬óįÕį┌ī”öĄŲ┌EPS«a┴┐║▄╔┘Ż¼Č°į┌ĘĆČ©Ų┌╗“╦ź£pŲ┌«a┴┐ät’@ų°į÷╝ė. SalimĄ╚▀Ć▀Mę╗▓ĮūCīŹEPSĄ─ĮM│╔īóļS╔·ķLų▄Ų┌Ą─▓╗═¼Č°ūā╗»(Salim et al., 2013). ę“┤╦Ż¼┼c╗Ņąį╬█─ÓŅÉ╦ŲŻ¼╬óįÕ╠Äė┌ĘĆČ©Ų┌╗“╦ź£pŲ┌Ģrūįą§─²ą¦╣¹▌^║├.į┌īŹļH┼ÓBųą┐╔īó╬óįÕĄ─╔·ķLļAČ╬┐žųŲį┌ĘĆČ©Ų┌╗“╦ź£pŲ┌ęį┤┘▀Mūį░ląįą§─²Ą─ą╬│╔.
ĪĪĪĪ4.4.4 Ąū╬’╦«ŲĮ
ĪĪĪĪNĪóPĄ╚ĀIBį¬╦žĄ─╚▒Ę”īó┤╠╝ż╬óįÕEPSĄ─╔·«aŻ¼▀@┼c╝ÜŠ·Ī󚵊·Ą╚╬ó╔·╬’ę╗ų┬. ╗∙ė┌┤╦Ż¼į┌▀\ąąųą┐╔▓╔ė├Ė▀├▄Č╚┼ÓBęį½@Ą├▌^Ą═Ą─F/MųĄŻ¼ęįūį╚╗ą╬│╔Ąū╬’╩▄Ž▐Ą─╣ż╦ćŚl╝■. Č°╬óįÕ╔·ķLĄ─┴Ēę╗ųžę¬Ąū╬’Ī¬Ī¬¤oÖC╠╝(IC)╩▄Ž▐ätīóęųųŲEPSĄ─╔·«a. ╚ńŻ¼Cordoba-CastroĄ╚░l¼Fą▒╔·¢┼įÕ(Scenedesmus obliquus)Ą─EPS╔·«aļSCO2Ą─╣®ĮoČ°į÷╝ėŻ║į┌Ė▀CO2╣®ĮoŚl╝■Ž┬(4%)Ż¼╬óįÕĄ─╔·ķL║═EPS«a┴┐Č╝ūŅ┤¾;Č°CO2╣®ĮoŽ┬ĮĄ║¾EPSĄ─╔·«aę▓ļSų«ĮĄĄ═.Å─ÅŖ╗»EPS╔·«aĄ─ĮŪČ╚üĒ┐┤Ż¼į┌īŹļH▀\ąąųą¤oę╔æ¬╝ėÅŖICĄ─╣®Įo. ╚╗Č°Ż¼╚ńPragyaĄ╚╦∙ųĖ│÷Ż¼×ķÅŖ╗»╗∙ė┌Ė▀pHĄ─ūįą§─²ät欎▐ųŲIC╣®ĮoŻ¼ęį▀_ĄĮ▒M┐╔─▄Ė▀Ą─pHŚl╝■.ę“┤╦Ż¼ī”ICĄ─š{┐žæ¬ÖÓ║ŌŲõī”Ė▀pH║═EPSā╔ĘNūįą§─²š²Ę┤ā╔ĘĮ├µĄ─ŠC║Žą¦æ¬.
ĪĪĪĪ4.4.5 ╬óįÕĘNī┘
ĪĪĪĪ¤ošō╩Ū╗∙ė┌Ė▀pHĄ─ūįą§─²▀Ć╩Ū╗∙ė┌EPSĄ─ūįą§─²Ż¼Ųõą§─²Śl╝■║═ą¦╣¹Č╝īóļS─┐ś╦įÕĘN▓╗═¼Č°«É(ģóęŖ4.2╣Ø║═4.3╣Ø▓┐Ęų). ▀@┐╔─▄╩Ūė╔ė┌įÕ╝Ü░¹į┌▒Ē├µ╠žąį║═╔·└Ē╠žąį╔ŽĄ─▓╗═¼Č°ī¦ų┬Ą─. ╚ńŻ¼ļŖžōąį▌^Ė▀Ą─įÕ╝Ü░¹ąĶę¬Ė³ČÓĦš²ļŖĄ─│┴ĄĒ╬’╔·│╔. ėų╚ńČÓ╝Ü░¹║═┤¾ą═ĮzĀŅįÕĘN▒╚å╬╝Ü░¹įÕĘNĖ³╚▌ęūą§─²│┴ĮĄ. į┌▀@ĘĮ├µąĶꬊC║Ž┐╝æ]╬óįÕĘNī┘Ą─╬█╦«ā¶╗»─▄┴”ĪóįÕ╝Ü░¹Ą─└¹ė├ārųĄĄ╚Ż¼▀xō±ąįĖ╗╝»ęūė┌ą§─²│┴ĮĄĄ─įÕĘN.
ĪĪĪĪ4.4.6 ╚▄ĮŌąįėąÖC╬’
ĪĪĪĪ┼c═Ō╝ė╗ņ─²ä®ŅÉ╦Ų(ģóęŖ3.1.2▓┐Ęų)Ż¼╦«ųą╚▄ĮŌąįėąÖC╬’(DOM)ī”ūįą§─²ę▓Ģ■«a╔·’@ų°Ą─ęųųŲū„ė├. ▀@ą®DOM╝╚┐╔─▄╩ŪįŁ╦«ųąÄ¦üĒĄ─Ė»ų│┘|Ż¼ę▓┐╔─▄╩ŪįÕŅÉ┤·ųx«a╔·Ą─ėąÖC╬’(AOM). BeuckelsĄ╚(2013)▒Ē├„Ż¼Ė»ų│╦ß║═įÕ╦ß¹}īó’@ų°ęųųŲ┴ū╦ßŌ}šTī¦Ą─ūįą§─²Ż¼Č°ŲŽ╠č╠Ū║═ęę╦ߥ╚ąĪĘųūėģsø]ėąė░Ēæ.═¼śėŻ¼WuĄ╚(2012)░l¼FAOMī”Üõč§╗»µVšTī¦Ą─ūįą§─²ėąÅŖ┴ęĄ─ęųųŲū„ė├Ż¼«öAOMÅ─┴Ńį÷╝ėų┴70 mg Īż L-1ĢrŻ¼ąĪŪ“įÕĄ─ūįą§─²ą¦┬╩Å─92%ĮĄĄ═ų┴7%.ŲõįŁę“║▄┐╔─▄╩ŪDOMīóā׎╚┼cCa2+ĪóMg2+Ą╚ļxūėĮY║ŽŻ¼Å─Č°ūĶų╣┴╦┴ū╦ßŌ}║═Üõč§╗»µVĄ╚│┴ĄĒ╬’Ą─«a╔·. ┴Ē═ŌŻ¼ė╔ė┌DOM▒Š╔ĒĦžōļŖŻ¼ę“┤╦Ģ■Ņ~═Ōį÷╝ėļŖąįųą║═╦∙ąĶĄ─ą§─²ä®ė├┴┐. Ķbė┌┤╦Ż¼BeuckelsĄ╚(2013)ųĖ│÷Ż¼DOMĄ─ęųųŲū„ė├║▄┐╔─▄╩Ū║▄ČÓīŹļHŪķørŽ┬Ż¼┴ū╦ßŌ}/Üõč§╗»µVĄ╚│┴ĄĒ╬’Ą─ŽÓĻP╔·│╔Śl╝■Č╝ęč│¼▀^┼RĮńųĄŻ¼Ą½ūįą§─²ģsø]ėą░l╔·Ą─įŁę“╦∙į┌.į┌▀@ĘĮ├µŲ╚ŪąąĶę¬Ė³▀Mę╗▓ĮĄ─ŽĄĮy蹊┐.
ĪĪĪĪ5 Ė„ĘNą§─²ĘųļxĘĮĘ©Ą─▒╚▌^┼cš╣═¹
ĪĪĪĪęįĶF¹}║═õX¹}×ķ┤·▒ĒĄ─Įī┘ą§─²ä®╩ŪĖ„ĘNą§─²ĘĮĘ©ųąæ¬ė├ūŅ×ķ│╔╩ņĄ─╝╝ąg.Ųõų„ę¬ā׳c╩Ū╦Ää®╔·«a║åå╬Ż¼ą§─²Śl╝■╚▌ęū┐žųŲŻ¼ą§─²ą¦╣¹ėą▒ŻšŽ. Ą½¤oÖCą§─²ä®Ą─ė├┴┐ę╗░Ń║▄┤¾(Äū░┘mg Īż L-1įÕę║)Ż¼Å─Č°«a╔·┤¾┴┐╬█─Ó. į┘š▀Ż¼ą§─²ą¦╣¹╩▄pHė░Ēæ▌^┤¾Ż¼ŲõūŅ╝čpHųĄ║▄┐╔─▄│¼│÷╬óįÕ┼ÓBŽĄĮyĄ─š²│ŻpHĘČć·Ż¼Ūę¤oÖCą§─²ä®āHī”▓┐Ęų╬óįÕĘNī┘ėąą¦. ūŅ▓╗└¹Ą─ą¦╣¹╩ŪŻ¼Įī┘¹}ŅÉ═∙═∙ī”įÕ╝Ü░¹Š▀ėąČŠ║”ū„ė├: ChenĄ╚į┌╩╣ė├┴“╦ßõX║═┬╚╗»ĶFą§─²¢┼įÕĢr░l¼FŻ¼«ö═Č╝ė┴┐▌^Ė▀ĢrŻ¼įÕ╝Ü░¹į┌24 h║¾╚½▓┐╦└═÷;PapaziĄ╚ę▓░l¼FõX¹}ļm╚╗ą§─²ą¦╣¹ūŅ║├Ż¼Ą½Ģ■ę²ŲįÕ╝Ü░¹┴čĮŌ.┤╦═ŌŻ¼Įī┘¹}ŅÉÜł┴¶į┌įÕ╝Ü░¹ųą▀Ćīóī”įÕ╝Ü░¹Ą─└¹ė├║═ūŅĮK╠Äų├įņ│╔▓╗└¹ė░Ēæ.ę“┤╦Ż¼Å─╬óįÕ┼ÓBĄ─ĮŪČ╚üĒ┐┤Ż¼Įī┘¹}ŅÉą§─²ä®▓ó▓╗╩ŪūŅ╝čĄ─╝╝ąg▀xō±. ėąĶbė┌┤╦Ż¼¤oÖCĮī┘ą§─²ä®╦Ų║§▓╗┐╔─▄│╔×ķ╬óįÕĘųļx▓╔╩šĄ─ų„ę¬░lš╣ĘĮŽ“.
ĪĪĪĪ┼c¤oÖCą§─²ä®ŽÓ▒╚Ż¼ėąÖCĖ▀Ęųūėą§─²ä®Š▀ėąĖ³Ė▀Ą─ą§─²ą¦┬╩(10Ī½30 mg Īż L-1įÕę║)Ż¼«a╔·Ą─╬█─Ó┴┐ąĪŻ¼─▄▀mė├ė┌Ė³ÅVĘ║Ą─╬óįÕĘNī┘.ŲõųąŻ¼Š█▒¹Ž®§Ż░Ęļm╚╗╩Ū╦«╠Ä└Ēųąæ¬ė├ūŅ│╔╩ņĄ─Ė▀Ęųūėą§─²ä®Ż¼Ą½Ųõī”╬óįÕĄ─ą§─²ą¦╣¹ģs▓╗╚ńÜżŠ█╠ŪĪóĻ¢ļxūėĄĒĘ█Ą╚╠ņ╚╗Ė▀Ęųūėą§─²ä®. ŪęŲõį┌╩╣ė├ųą┐╔─▄Ģ■ßīĘ┼│÷ę╗Č©┴┐Š▀ėąÅŖ┴ęČŠąįĄ─å╬¾w▒¹Ž®§Ż░ĘŻ¼ę“┤╦Ųõæ¬ė├Ū░Š░ėąŽ▐. ╠ņ╚╗Ė▀Ęųūėą§─²ä®¤oČŠŻ¼ęū╔·╬’ĮĄĮŌŻ¼ī”╬óįÕ┼ÓB║═įÕ╝Ü░¹Ą─║¾└m└¹ė├╗∙▒Š¤ožōū„ė├Ż¼į┌╬óįÕĄ─Ęųļx▓╔╩šųąŠ▀ėą┴╝║├Ą─æ¬ė├Øō┴”. Ą½╠ņ╚╗Ė▀Š█╬’ųąų╗ėąÜżŠ█╠ŪĄ╚╔┘öĄ╩ŪĻ¢ļxūėą═Ą─. ÜżŠ█╠ŪĄ─ą§─²ą¦┬╩║▄Ė▀Ż¼Ą½Ųõą§─²Śl╝■ę╗░Ń×ķ╦ßąįŻ¼│¼│÷┴╦╬óįÕ╔·ķLĄ─š²│ŻpHĘČć·. ┐╝æ]ĄĮī”┤¾┴┐įÕę║▀Mąą╦ß╗»╦∙ąĶ═Č╝ėĄ─╗»īW╦Ää®ė├┴┐Ż¼ÜżŠ█╠Ū║▄┐╔─▄į┌ĮøØ·╔Ž▓╗Š▀éõ▀xō±ąį. Ļ¢ļxūėĄĒĘ█į┌įŁ┴Ž╔Ž┐╔┤¾┴┐½@╚ĪŻ¼ārĖ±Ą═┴«Ż¼═Č╝ė┴┐ĘŪ│ŻąĪ(Äūmg Īż L-1įÕę║)Ż¼ą§─²ą¦╣¹ā׫ÉŪę╗∙▒Š▓╗╩▄pHė░ĒæŻ¼Š▀ėą┴╝║├Ą─╣ż│╠╗»æ¬ė├Øō┴”. ║¾└m蹊┐Ą─ųž³cæ¬į┌ė┌ā×╗»ŲõĻ¢ļxūė╗»▀^│╠Ż¼ęį▀Mę╗▓Į╠ßĖ▀Ųõ▀mė├ąį║═ą§─²ą¦┬╩Ż¼▓ó’@ų°ĮĄĄ═╝ė╣żųŲįņ│╔▒Š.
ĪĪĪĪ└¹ė├╝ÜŠ·Ī󚵊·Ą╚╬ó╔·╬’╔·«aĄ─╔·╬’ą§─²ä®Š▀ėąĖ▀ą¦Īó¤oČŠĪó┐╔╔·╬’ĮĄĮŌĄ╚ā׳c. Ą½ŲõĖ„ĘN└¹ė├ĘĮ╩ĮČ╝┤µį┌├„’@╚▒Ž▌Ż║1)ų▒Įė═Č╝ė╬ó╔·╬’╝Ü░¹╗“Š·-įÕ╣▓═¼┼ÓBėąī”╬óįÕ┼ÓBŽĄĮyįņ│╔╬█╚ŠĄ─’LļU;2)═Č╝ė┼ÓBę║Īó│ķ╚Īę║Īó╠ß╚Ī╬’Ą╚ĘĮ╩ĮąĶę¬ę╗éĆ╬óįÕ┼ÓBŽĄĮyęį═ŌĄ─å╬¬Ü┼ÓB¾wŽĄŻ¼ė╚Ųõ╩Ū║¾ā╔š▀▀Ć╔µ╝░ĄĮÅ═ļsĄ─Ęųļx║═╝ė╣żå¢Ņ}Ż¼▀@¤oę╔Ģ■į÷╝ė└¹ė├ļyČ╚║═│╔▒Š. ┐╔ęŖŻ¼╔·╬’ą§─²ä®ę╗░Ń╦∙ą¹ĘQĄ─Ą═│╔▒Šā×ä▌┐╔─▄į┌īŹ█`ųąļyęį─▄│╔×ķ¼FīŹ. ╔·╬’ą§─²ä®┴Ēę╗╚▒³c╩ŪŻ¼─│ę╗╔·╬’ą§─²ä®┐╔─▄ų╗ī”─│ą®╠žČ©įÕĘNą§─²ą¦╣¹▌^║├. └¹ė├ūįą§─²įÕĘN«a╔·Ą─╔·╬’ą§─²ä®▓╗ąĶę¬Ņ~═ŌĄ─┼ÓB¾wŽĄŻ¼Ūę¤o╬█╚Š╬óįÕ┼ÓBų«ė▌. Ą½ūįą§─²ĘNī┘Ą─╔·ķL╦┘Č╚ę╗░ŃĄ═ė┌ĘŪūįą§─²ĘNī┘Ż¼Ųõ╬█╦«ā¶╗»─▄┴”║═«aė═Øō┴”ę▓┐╔─▄▓╗╚ńĘŪą§─²įÕĘN.ę“┤╦Ż¼┐žųŲūįą§─²įÕĘNį┌ŽĄĮyųąĄ─▒╚└²ų┴ĻPųžę¬. ▀@Š═╠ß│÷┴╦į┌╗ņ║Ž┼ÓBųą▀MąąĘN╚║┐žųŲĄ─Å═ļsę¬Ū¾. ┼c╝ÜŠ·Ī󚵊·Ą╚╬ó╔·╬’«a╔·Ą─╔·╬’ą§─²ä®ŅÉ╦ŲŻ¼ūįą§─²įÕųĻĄ─ą§─²ą¦╣¹ę▓īóļS─┐ś╦įÕĘNĄ─▓╗═¼Č°«É. └ĒĮŌūįą§─²(EPSšTī¦)įÕĘNĄ─ą§─²ÖC└Ēī”┤┘▀MŲõæ¬ė├Š▀ėąĻPµIęŌ┴x. ─┐Ū░į┌▀@ĘĮ├µĄ─蹊┐▀ĆĘŪ│Ż▓╗ūŃŻ¼╗∙▒Š╔Ž▀Ć╠Äė┌ī”EPSĄ─│╔ĘųĘų╬÷╔Ž. DCB└Ēšōļm╚╗─▄ĮŌßī║▄ČÓīŹ“×¼FŽ¾Ż¼Ą½┤µį┌▓╗─▄ĮŌßī×ķ╩▓├┤ų╗ėą╠žČ©įÕĘN▓┼Š▀ėąą§─²ū„ė├▀@ę╗Ė∙▒Š╚▒Ž▌. į┌┤╦ĘĮ├µŻ¼ė╔įÕ╝Ü░¹ėH/╩Ķ╦«ąįøQČ©Ą─Lewis╦ß-ēA╦«║Žū„ė├┴”╩ŪĘŪ│ŻėąŪ░Š░Ą─└ĒšōŻ¼æ¬įō│╔×ķ║¾└m蹊┐Ą─ųž³c.
ĪĪĪĪÜõč§╗»µV│┴ĄĒļm╚╗─▄ėąą¦šTī¦│÷ūį░ląįą§─²Ż¼Ą½Ųõą╬│╔ę╗░Ńę¬į┌pH>10.5ęį╔Ž. Č°┤¾▓┐Ęų╬óįÕį┌pH>9Ģr╣Ō║Žū„ė├Š═Ģ■╩▄ĄĮ’@ų°ęųųŲ╔§ų┴═Ļ╚½═Żų╣Ż¼╦∙ęį╬óįÕĄ─ūį╚╗╔·ķL║▄┐╔─▄▀_▓╗ĄĮÜõč§╗»µV│┴ĄĒĄ─╔·│╔Śl╝■. ╩┬īŹ╔ŽŻ¼Äū║§╦∙ėą╗∙ė┌Üõč§╗»µV│┴ĄĒĄ─ūįą§─²Č╝╩Ū═©▀^═Ō╝ėēAąį╬’┘|▀_ĄĮ╦∙ąĶpHųĄ(▒Ē 6)Ż¼▀@¤oę╔Ģ■ĦüĒŅ~═ŌĄ─│╔▒Š. Ė▀pH▀Ć┐╔─▄ī”įÕ╝Ü░¹įņ│╔ć└ųžōpé¹Ż¼└²╚ńŻ¼Ė▀«aė═įÕĘNSkeletoma costatumį┌pH = 10. 2Ģrą§─²┬╩▀_ 80%Ż¼Ą½╗ž╩šįÕ¾wųąŽÓ«öę╗▓┐Ęų╝Ü░¹░l╔·ĮŌ¾w╝░░¹ā╚│╔Ęų═Ōą╣Ż¼ć└ųžė░Ēæ║¾└mČ■╩«╠╝╬ÕŽ®╦ß(EPA)╠ß╚Ī╣ż╦ć;┬▌ą²įÕ(Spirulina platensis)į┌pHĖ▀ė┌13ĢrŻ¼╝Ü░¹ą§─²Ą├ėų┐ņėųÅžĄūŻ¼Ą½┤╦ĢrįÕ╝Ü░¹Ņü╔½░l³SŻ¼▒Ē├„Ųõ╝Ü░¹ęč╩▄ĄĮ▌^×ķć└ųžĄ─Ī░pHōpé¹Ī▒.┤╦═ŌŻ¼Ė▀pHųĄ▀Ć║▄┐╔─▄│¼│÷┼┼Ę┼ś╦£╩Ż¼ąĶę¬į┘╝ė╦ßš{╣ØĄĮ╚▌įSĄ─ĘČć·. ę“┤╦Ż¼╗∙ė┌Üõč§╗»µV│┴ĄĒĄ─ūįą§─²ų╗▀mė├ė┌å╬╝āĄ─╬óįÕ▓╔╩šŻ¼ī”╬óįÕ╬█╦«╠Ä└ĒŽĄĮyüĒšf▓ó▓╗╩Ūę╗éĆ║Ž▀mĄ─▀xō±.
ĪĪĪĪ╗∙ė┌┴ū╦ßŌ}│┴ĄĒĄ─ūįą§─²¤oąĶ╚╬║╬Ņ~═Ō═Č╚ļŻ¼į┌╬óįÕūį╚╗╔·ķLĄ─pHĘČć·ā╚(8Ī½10)Š═─▄ą╬│╔Ż¼─▄═¼▓ĮīŹ¼F│²┴ūŻ¼ī”╬óįÕ╗Ņąį║═įÕ╝Ü░¹Ą─║¾└m╝ė╣ż└¹ė├Äū║§ø]ėą▓╗└¹ė░Ēæ. ę“┤╦Ż¼¤ošō╩ŪÅ─╬█╦«╔ŅČ╚╠Ä└ĒŻ¼▀Ć╩ŪÅ─įÕ╝Ü░¹Ą─▓╔╩š└¹ė├Ą╚ĮŪČ╚üĒ┐┤Č╝╩ŪūŅ║Ž▀mĄ─Ęųļx▓╔╩šĘĮĘ©ų«ę╗. ė╚Ųõ╩ŪļSų°┴ū╦ß¹}ØŌČ╚╠ßĖ▀Ż¼Ųõ┼RĮńpHųĄīó’@ų°Ž┬ĮĄ;Č°▀@į┌īŹļH╬█╦«╠Ä└ĒųąŪĪŪĪ╩Ūę╗éĆ║▄╚▌ęū┐žųŲĄ─╣ż╦ćŚl╝■. Ķbė┌┤╦Ż¼┐╔ęį╠ß│÷ęįŽ┬ÄūéĆÅŖ╗»╗∙ė┌┴ū╦ßŌ}│┴ĄĒūįą§─²Ą─╦╝┬ĘŻ║ó┘į┌╬█╦«╠Ä└ĒĄ─ų„¾w╣ż╦ćųą╚ĪŽ¹ÅŖ╗»╔·╬’│²┴ūŻ¼×ķ║¾└m╬óįÕ╠Ä└ĒŽĄĮy▒Ż┴¶Ė▀┴ūØŌČ╚;ó┌▓╔ė├Ė▀├▄Č╚ķgą¬┼ÓBĘĮ╩ĮŻ¼▓ó═Ļ╚½╗“į┌Ę┤æ¬ų▄Ų┌─®Č╦╚ĪŽ¹═Ō▓┐CO2╣®ĮoŻ¼ęįčĖ╦┘Ūęūį╚╗Ąžą╬│╔Ė▀pHŚl╝■;ó█═©▀^▀m«öčėķLĘ┤æ¬ų▄Ų┌ĪóÅŖ╗»╣Ō║Žū„ė├(╣ŌššĪóĄū╬’ØŌČ╚)Ą╚▀Mę╗▓Į┤┘▀MpHĄ─╠ß╔². ─┐Ū░┤µį┌Ą─å¢Ņ}╩Ū┤¾▓┐Ęų蹊┐Č╝ų╗╩Ū┐╝▓ņĖ▀pHŽ┬╦▓ĢrĄ─ą§─²ą¦╣¹. ╚ń╣¹╬óįÕ┼ÓBŽĄĮyķLŲ┌╠Äė┌šTī¦ūįą§─²╦∙ąĶĄ─Ė▀pHŁhŠ│Ż¼ÄūéĆĘŪ│ŻųĄĄ├ĻPūóĄ─å¢Ņ}╩ŪŻ║ó┘╬óįÕĘN╚║ĮYśŗ╩ŪʱĢ■░l╔·’@ų°ūā╗»?ó┌ ─┐ś╦įÕĘN─▄ʱŠS│ųā×ä▌?ó█ ╬óįÕĄ─╔·└Ē╠žąį(ā¶╗»─▄┴”Īóė═ų¼║¼┴┐Ą╚)╩ŪʱĢ■░l╔·Ė─ūā?▀@ą®Č╝ąĶę¬▀Mę╗▓ĮĄ─ŽĄĮy蹊┐üĒ├„┤_.
ĪĪĪĪ6 ĮYšō
ĪĪĪĪė├ė┌╬óįÕĘųļxĪó▓╔╩šĄ─└ĒŽļą§─²ä®æ¬įōŠ▀ėą¤oČŠ(ī”įÕ╝Ü░¹▒Š╔Ē╝░ŁhŠ│)ĪóĄ═│╔▒ŠĪóÅVūVĖ▀ą¦Īó▓╗ė░ĒæįÕ╝Ü░¹║¾└m└¹ė├Ą╚╠ž³c. ę└┤╦ś╦£╩Ż¼Įī┘¹}ŅÉą§─²ä®ę“═Č╝ė┴┐┤¾Īóī”įÕ╝Ü░¹ėąČŠąį╝░ė░ĒæįÕ╝Ü░¹Ą─║¾└m└¹ė├Ą╚╚▒³c▓╗─▄│╔×ķ╬óįÕą§─²Ą─ų„ę¬░lš╣ĘĮŽ“. ėąÖCĖ▀Ęųūėą§─²ä®ųąŠ█▒¹Ž®§Ż░Ęī”╬óįÕĄ─ą§─²ą¦╣¹▌^▓ŅŻ¼┐╔─▄ßīĘ┼ėąČŠĄ─▒¹Ž®§Ż░Ęå╬¾wŻ¼ę“┤╦Ųõæ¬ė├Ū░Š░ėąŽ▐. ÜżŠ█╠Ūą§─²ą¦┬╩▌^Ė▀Ż¼Ą½▀mæ¬Ą─ą§─²pHę╗░Ń×ķ╦ßąįŻ¼│¼│÷┴╦╬óįÕ┼ÓBĄ─š²│ŻĘČć·Ż¼Š▀ėą║▄┤¾Ą─ŠųŽ▐ąį. Č°Ļ¢ļxūėĄĒĘ█į┌įŁ┴Ž╔Ž┐╔┤¾┴┐½@╚ĪŻ¼ārĖ±Ą═┴«Ż¼═Č╝ė┴┐ĘŪ│ŻąĪŻ¼ą§─²ą¦╣¹ā׫ÉŪę╗∙▒Š▓╗╩▄pHė░ĒæŻ¼Š▀ėą┴╝║├Ą─╣ż│╠╗»æ¬ė├Øō┴”. ║¾└m蹊┐Ą─ųž³cæ¬į┌ė┌ā×╗»ŲõĻ¢ļxūė╗»▀^│╠Ż¼ęį▀Mę╗▓Į╠ßĖ▀Ųõ▀mė├ąį║═ą§─²ą¦┬╩Ż¼▓ó’@ų°ĮĄĄ═╝ė╣żųŲįņ│╔▒Š.
ĪĪĪĪ└¹ė├╝ÜŠ·Ī󚵊·Ą╚╬ó╔·╬’╔·«aĄ─╔·╬’ą§─²ä®Š▀ėąĖ▀ą¦Īó¤oČŠĪó┐╔╔·╬’ĮĄĮŌĄ╚ā׳c. Ą½ŲõĖ„ĘN└¹ė├ĘĮ╩ĮČ╝┤µį┌├„’@╚▒Ž▌Ż¼└²╚ńŻ¼┐╔─▄ī”╬óįÕ┼ÓBįņ│╔╬█╚ŠŻ¼ąĶę¬Ņ~═ŌĄ─┼ÓBŽĄĮyŻ¼╔µ╝░ĄĮÅ═ļsĄ─Ęųļx╝ā╗»▀^│╠Ą╚. ┤╦═ŌŻ¼╔·╬’ą§─²ä®Ą─ą§─²ÖC└Ē║═Śl╝■Č╝▀Ć▓╗╔§├„┤_. ę“┤╦Ż¼╔·╬’ą§─²ä®ŠÓ╣ż│╠╗»æ¬ė├▀Ć┤µį┌║▄┤¾ŠÓļx. ═¼└ĒŻ¼╗∙ė┌░¹═ŌŠ█║Ž╬’Ą─╬óįÕūįą§─²ę“ą§─²ÖC└ĒÅ═ļsĪóą§─²Śl╝■▓╗├„┤_╝░╔µ╝░ĄĮÅ═ļsĄ─ĘN╚║┐žųŲę¬Ū¾Ą╚Ż¼┐╔┐┐ąį▌^▓ŅŻ¼į┌╬óįÕą§─²Ęųļxųą┐╔─▄ų╗─▄ŲĄĮÕ\╔Ž╠Ē╗©Ą─ū„ė├.
Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.bnynw.comĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻĪĪĪĪ╗∙ė┌Üõč§╗»µV│┴ĄĒĄ─ūįą§─²ę¬Ū¾║▄Ė▀Ą─pHųĄ(>10.5). ▀@═∙═∙│¼│÷╬óįÕš²│Ż╔·ķL╦∙─▄▀_ĄĮĄ─pHųĄĘČć·Ż¼ąĶę¬Ņ~═Ō═Č╝ėēAąį╬’┘|Ż¼┐╔─▄ī”įÕ╝Ü░¹Ą─╗Ņąį║═║¾└m└¹ė├įņ│╔▓╗└¹ė░ĒæŻ¼ę“┤╦Ż¼ī”╬óįÕ╬█╦«╠Ä└ĒŽĄĮy┐╔─▄▓ó▓╗╩Ū║Ž▀mĄ─▀xō±. ╗∙ė┌┴ū╦ßŌ}│┴ĄĒĄ─ūįą§─²¤oąĶ╚╬║╬Ņ~═Ō═Č╚ļŻ¼į┌╬óįÕūį╚╗╔·ķLĄ─pHĘČć·ā╚(8Ī½10)Š═─▄ą╬│╔Ż¼─▄═¼▓ĮīŹ¼F╗»īW│²┴ūŻ¼ī”╬óįÕ╗Ņąį║═įÕ╝Ü░¹Ą─║¾└m╝ė╣ż└¹ė├Äū║§ø]ėą▓╗└¹ė░Ēæ. ę“┤╦Ż¼¤ošō╩ŪÅ─╬█╦«╔ŅČ╚╠Ä└ĒŻ¼▀Ć╩ŪÅ─įÕ╝Ü░¹Ą─▓╔╩š└¹ė├Ą╚ĮŪČ╚üĒ┐┤Č╝╩ŪūŅ║Ž▀mĄ─Ęųļx▓╔╩šĘĮĘ©ų«ę╗. ║¾└m蹊┐æ¬▀Mę╗▓Į├„┤_Ųõą╬│╔Śl╝■Ż¼▓óĮY║Ž╬█╦«╠Ä└Ēų„¾w╣ż╦ćĄ─š{š¹(╚ĪŽ¹ÅŖ╗»│²┴ū)Īó╬óįÕ┼ÓBĘĮ╩ĮĄ─Ė─▀M(Ė▀├▄Č╚ķgą¬┼ÓBĪó╚ĪŽ¹CO2╣®ĮoŻ¼š{╣ØĘ┤æ¬ų▄Ų┌)Ą╚ī”Ųõ▀MąąÅŖ╗». ┤╦═ŌŻ¼▀Ćæ¬ųž³cĻPūó╬óįÕ┼ÓBŽĄĮyķLŲ┌╠Äė┌Ė▀pHŚl╝■Ž┬┐╔─▄░l╔·Ą─ĘN╚║ĮYśŗ║═╔·└Ē╠žąįūā╗».

